Key Objectives
By the end of this section, you will be able to do the following:
- Describe the structure of prokaryotic and eukaryotic genomes
- Distinguish between chromosomes, genes, and traits
- Describe the mechanisms of chromosome compaction
The continuity of life from one cell to another has its foundation in the reproduction of cells by way of the cell cycle. The cell cycle is an orderly sequence of events that describes the stages of a cell’s life from the division of a single parent cell to the production of two new genetically identical daughter cells.
Genomic DNA
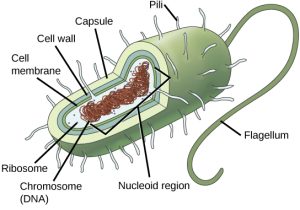
Before discussing the steps a cell must undertake to replicate and divide its DNA, a deeper understanding of the structure and function of a cell’s genetic information is necessary. A cell’s DNA, packaged as a double-stranded DNA molecule, is called its genome. In prokaryotes, the genome is composed of a single, double-stranded DNA molecule in the form of a loop or circle (Figure 10.2). The region in the cell containing this genetic material is called a nucleoid. Some prokaryotes also have smaller loops of DNA called plasmids that are not essential for normal growth. Bacteria can exchange these plasmids with other bacteria, sometimes receiving beneficial new genes that the recipient can add to their chromosomal DNA. Antibiotic resistance is one trait that often spreads through a bacterial colony through plasmid exchange from resistant donors to recipient cells.
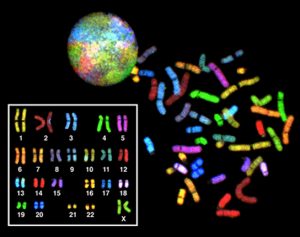
In eukaryotes, the genome consists of several double-stranded linear DNA molecules (Figure 10.3). Each species of eukaryotes has a characteristic number of chromosomes in the nuclei of its cells. Human body (somatic) cells have 46 chromosomes, while human gametes (sperm or eggs) have 23 chromosomes each. A typical body cell contains two matched or homologous sets of chromosomes (one set from each biological parent)—a configuration known as diploid. (Note: The letter n is used to represent a single set of chromosomes; therefore, a diploid organism is designated 2n.) Human cells that contain one set of chromosomes are called gametes, or sex cells; these are eggs and sperm, and are designated 1n, or haploid.
Upon fertilization, each gamete contributes one set of chromosomes, creating a diploid cell containing matched pairs of chromosomes called homologous (“same knowledge”) chromosomes. Homologous chromosomes are the same length and have specific nucleotide segments called genes in exactly the same location, or locus. Genes, the functional units of chromosomes, determine specific characteristics by coding for specific proteins. Traits are the variations of those characteristics. For example, hair color is a characteristic with traits that are blonde, brown, or black, and many colors in between.
Apparently minor variations of traits, such as blood type, eye color, and handedness, contribute to the natural variation found within a species, but even though they seem minor, these traits may be connected with the expression of other traits as of yet unknown. However, if the entire DNA sequence from any pair of human homologous chromosomes is compared, the difference is much less than one percent. The sex chromosomes, X and Y, are the single exception to the rule of homologous chromosome uniformity: Other than a small amount of homology that is necessary to accurately produce gametes, the genes found on the X and Y chromosomes are different.
Eukaryotic Chromosomal Structure and Compaction
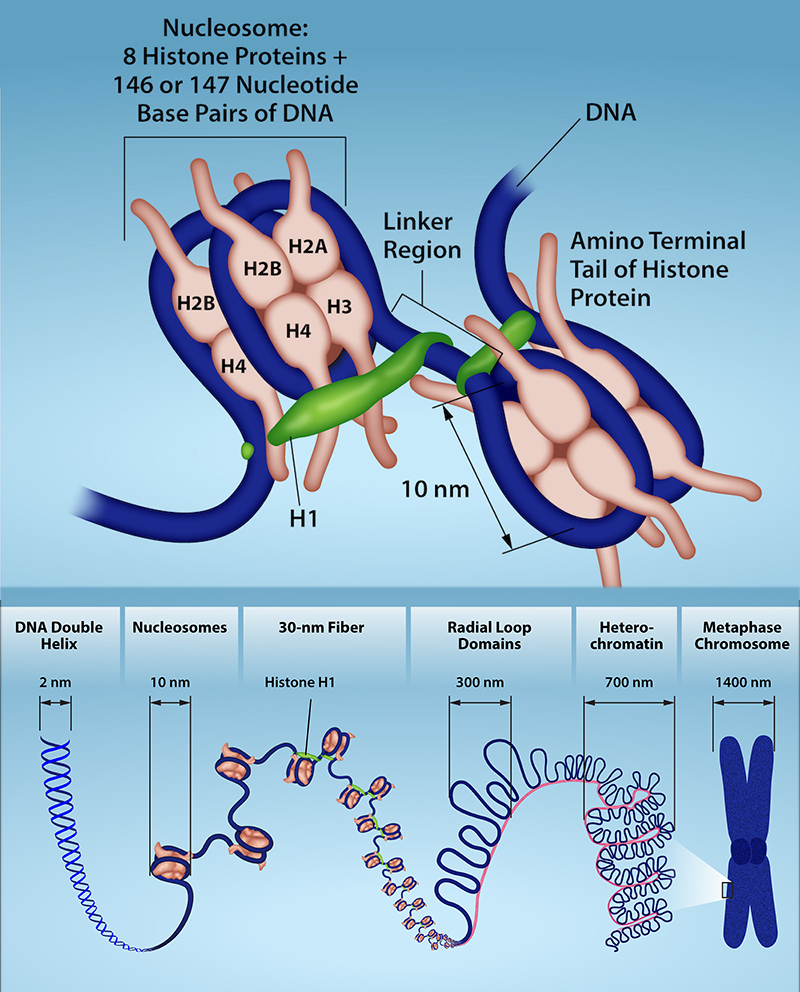
If the DNA from all 46 chromosomes in a human cell nucleus were laid out end-to-end, it would measure approximately two meters; however, its diameter would be only 2 nm! Considering that the size of a typical human cell is about 10 µm (100,000 cells lined up to equal one meter), DNA must be tightly packaged to fit in the cell’s nucleus. At the same time, it must also be readily accessible for the genes to be expressed. For this reason, the long strands of DNA are condensed into compact chromosomes during certain stages of the cell cycle. There are a number of ways that chromosomes are compacted.
In the first level of compaction, short stretches of the DNA double helix wrap around a core of eight histone proteins at regular intervals along the entire length of the chromosome (Figure 10.4). The DNA-histone complex is called chromatin. The beadlike, histone DNA complex is called a nucleosome, and DNA connecting the nucleosomes is called linker DNA. A DNA molecule in this form is about seven times shorter than the double helix without the histones, and the beads are about 10 nm in diameter, in contrast with the 2-nm diameter of a DNA double helix.
The second level of compaction occurs as the nucleosomes and the linker DNA between them coil into a 30-nm chromatin fiber. This coiling further condenses the chromosome so that it is now about 50 times shorter than the extended form.
In the third level of compaction, a variety of fibrous proteins is used to “pack the chromatin.” These fibrous proteins also ensure that each chromosome in a non-dividing cell occupies a particular area of the nucleus that does not overlap with that of any other chromosome (see the top image in Figure 10.3).
DNA replicates in the S phase of interphase, which technically is not a part of mitosis, but must always precede it. After replication, the chromosomes are composed of two linked sister chromatids. When fully compact, the pairs of identically packed chromosomes are bound to each other by cohesin proteins. The connection between the sister chromatids is closest in a region called the centromere. The conjoined sister chromatids, with a diameter of about 1 µm, are visible under a light microscope. The centromeric region is highly condensed and thus will appear as a constricted area.
Link to Learning
This animation illustrates the different levels of chromosome packing.
