5.2 Selective Permeability and Solute Gradients
KEY CONCEPTS
By the end of this section, you will be able to do the following:
- Evaluate how the components of membranes that impact the selective permeability of those membranes to different types of solutes (small/large, hydrophobic/hydrophilic, neutral/charged).
- Compare and contrast how channel proteins and carrier proteins facilitate solute movement across membranes.
- Explain how concentration gradients and electrochemical gradients impact the movement of different types of solutes across membranes.
All cells and organelles need some kind of physical barrier to keep their contents in and external materials out. They also need a way of exchanging materials between their internal and external environments. This section explores how selectively permeable membranes accomplish this, and how solute gradients influence the movement of materials across these membranes.
Selective Permeability
Membranes must allow certain substances to enter or leave while preventing other substances from passing though. In other words, plasma membranes are selectively permeable. If plasma membranes were to lose this selectivity, the cell would no longer be able to sustain itself and would die. In addition to the plasma membrane, most organelles are surrounded by their own membranes, and these membranes are also selectively permeable. Whether a solute can cross a membrane depends on at least two things: the properties of the solute and the properties of the membrane itself.
Solute Properties
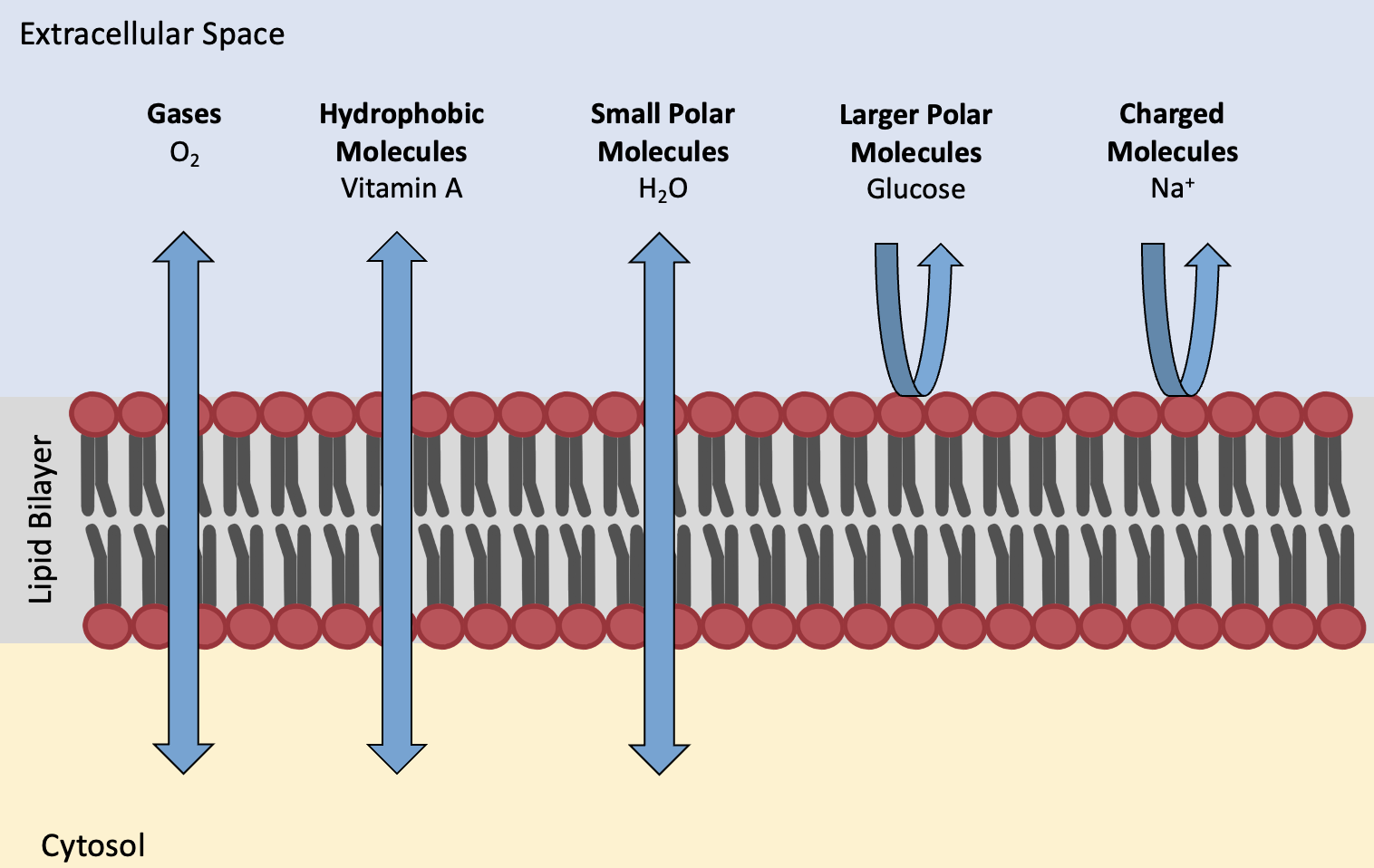
A solute is a substance that can be dissolved in a solvent. Recall that membrane lipids are amphiphilic: they have hydrophilic and hydrophobic regions, resulting in the lipid bilayer having a hydrophobic core. This characteristic helps move some materials through the membrane and hinders the movement of others. Gasses, hydrophobic molecules, and small polar (hydrophilic) molecules like water can easily slip through the membrane’s hydrophobic lipid core. Some examples of solutes that easily enter or exit human cells include lipid-soluble (hydrophobic) vitamins A, D, E, and K lipid-soluble drugs and hormones, and oxygen and carbon dioxide molecules. Water is not lipid-soluble, but its small size allows it to diffuse across membranes anyway (Figure 5.14).
Some solutes cannot readily pass through membranes. For instance, polar substances cannot easily diffuse through the membrane’s non-polar lipid core (except for very small molecules like water). Ions are charged, which prevents them from directly crossing membranes. Ions of hydrogen (H+), sodium (Na+), potassium (K+), calcium (Ca2+), chloride (Cl–), and many others must enter or exit cells with the help of transmembrane proteins (channels/carriers). Larger polar molecules, such as simple sugars and amino acids, also need the help of various transmembrane proteins (channels/carriers) to transport themselves across membranes. Some molecules, such as proteins and polysaccharides like glycogen, are too large to cross the membrane at all (Figure 5.14). These molecules must be moved across the membrane via vesicular transport, which is discussed further in Chapter 5.5.
Membrane Properties
The permeability of membranes is affected by a number of factors. First, the permeability of the membrane is related to its fluidity. The more fluid the membrane, the more permeable it is to substances that can pass through it without the aid of a transport protein. For polar and charged substances that do require a transport protein to cross the membrane, the transport proteins in the membrane greatly affect its permeability. The more transport proteins there are in the membrane, the more permeable the membrane to the specific solute that uses that transport protein. Similarly, the higher the proportion of transport proteins that are active/open, the higher the permeability. There are two categories of transport proteins that can facilitate solute movement across membranes: channel proteins and carrier proteins. Understanding these types of proteins is important for understanding membrane permeability, and will also help with your understanding of passive and active transport in Chapters 5.3 and 5.4.
Channel proteins are transmembrane proteins with a hydrophilic channel through their core that provides an opening through the membrane (Figure 5.15). This passage through the channel allows specific charged or polar solutes to avoid the membrane’s hydrophobic central portion that would otherwise slow or prevent their entry into the cell. Each type of channel is specific for the transported solute. For example, aquaporins are channel proteins that allow water (but not most other solutes) to pass through the membrane at a very high rate. If a cell has aquaporins in its plasma membrane, that plasma membrane has higher permeability to water than a cell lacking aquaporins.
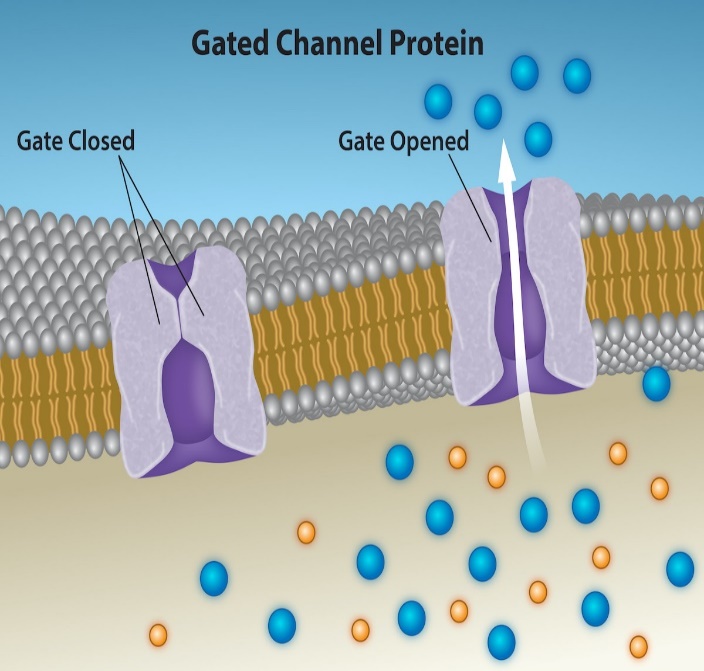
Some channel proteins are open at all times (e.g., aquaporins), while others are “gated” (Figure 5.15, in which case the protein can change its conformation to block the channel’s opening. In some cell types, sodium and chloride ions pass freely through open channels, whereas in other cell types a gate must open to allow passage. A cell with open sodium ion channels has higher permeability to sodium ions than a cell with closed sodium ion channels (or no sodium ion channels). Animal cells involved in transmitting electrical impulses, such as neuron and muscle cells, have gated channels for sodium, potassium, and calcium in their membranes. Opening and closing these channels allows ions to cross the membrane, changing the relative concentrations of these ions on opposing sides of the membrane of these ions, which facilitates electrical impulse transmission along membranes (in the case of neurons) or muscle contraction (in the case of muscle cells).
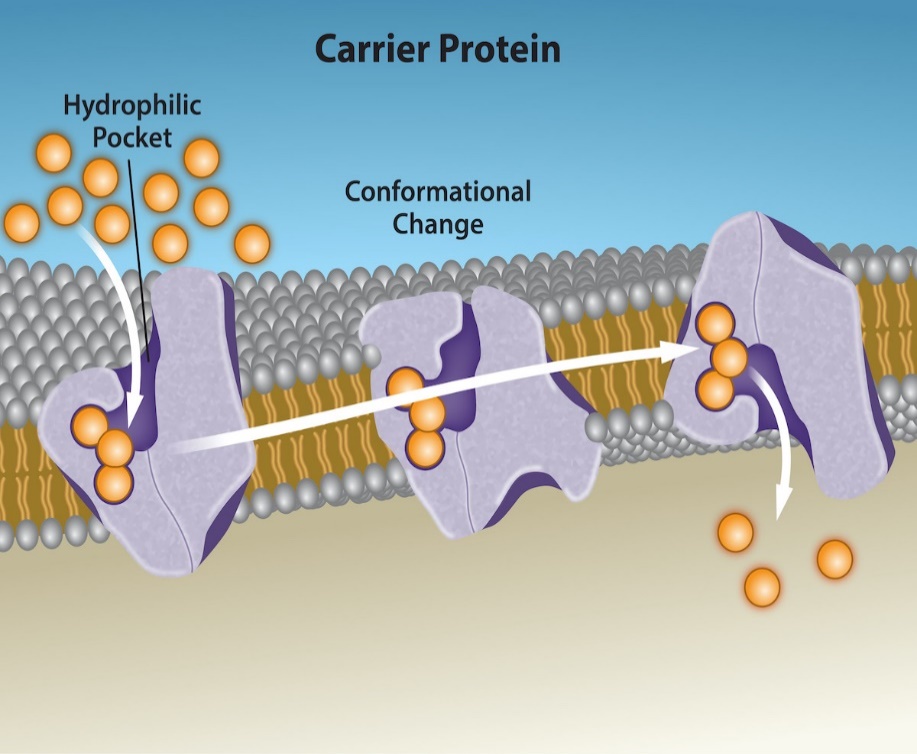
Carrier proteins bind a specific solute and then change shape to move the bound solute into or out of the cell (Figure 5.16). Like channels, carrier proteins are typically specific for a single solute. For example, glucose transporters (GLUTs) are a group of carrier proteins that facilitate the transport of glucose across the membrane. Binding of glucose to the binding site on the carrier protein induces a conformational change associated with transport, and releases glucose to the other side of the membrane. Some animal cell types (e.g., human liver cells) contain increase the abundance of GLUT transporters in the plasma membrane in order to increase membrane permeability to glucose when blood sugar levels are high, allowing that glucose to leave the blood and enter cells.
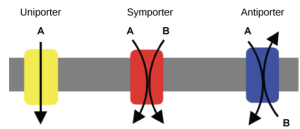
Carrier proteins can be characterized by how many different solutes they move, and in what direction those solutes are transported. Some common forms of carrier proteins include uniporter, symporter and antiporter proteins (Figure 5.17). Uniporter proteins facilitates the movement of a single type of solute across the membrane. For example, glucose transporters (GLUT) are a form of uniporter proteins that help regulate blood glucose levels by allowing blood glucose to enter cells within animal bodies. Symport (co-transport) proteins facilitate simultaneous transport of two or more different solutes in the same direction across a membrane. For example, some of the cells lining animal intestines contain sodium-glucose symporters, which move both sodium ions and glucose molecules from the intestine (gut) into the cells. Antiport (counter-transport) proteins facilitate simultaneous transport of two or more different solutes in opposite directions across a membrane. For example, most animal cells have Na+/K+ ATPase transporters in the plasma membrane; these are sodium-potassium antiporters that move three sodium ions out of the cell and two potassium ions into the cell with each movement of the carrier protein. We will see additional examples and explanations of uniporter, symporter, and antiporter proteins in Chapters 5.3 and 5.4.
While channel and carrier proteins fulfil similar functions, they operate at different rates. Channel proteins facilitate solute movement across membranes at a rate of tens of millions of solutes per second. Conversely, carrier proteins are much slower, working at a rate of a thousand to a million solutes per second. In the latter case, solute movement is slower because a carrier protein must change shape to move each solute (usually one at a time) across the membrane.
Solute Gradients
Some cells require larger amounts of specific solutes and must have a way of obtaining these materials from extracellular fluids. Some solutes are so important to a cell that it spends some of its energy by hydrolyzing adenosine triphosphate (ATP) to obtain these materials. For example, most of a red blood cell’s energy is used to maintain internal sodium and potassium levels that differ from those of the surrounding environment. Whether a cell needs to expend energy to transport a solute into or out of the cell depends on the gradient of that solute across the membrane. There are two types of relevant gradients that we will discuss: concentration gradients and electrochemical gradients. A strong understanding of gradients is important for understanding the content in Chapters 5.3 and 5.4.
Concentration Gradients (uncharged solutes)
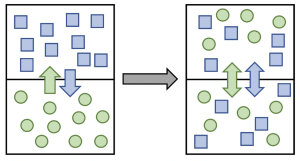
A concentration gradient occurs when the concentration of something (e.g., a type of solute) is higher in one area than another, for example inside versus outside the cell. When thinking about this in the context of membrane permeability, concentration gradients are most relevant for uncharged (neutral) solutes which may be hydrophilic (e.g., glucose) or hydrophobic (e.g., oxygen). When a concentration gradient of a solute exists across a membrane, and that membrane is permeable to the solute, there will be a net movement of solute from the area of higher concentration to the area of lower concentration (down or with the concentration gradient), until the concentration of that solute is equal on either side of the membrane. When there is no longer a concentration gradient, we say that the solute has reached dynamic equilibrium, a state in which there is no net movement of solute across the membrane. That is, solutes can still move across the membrane in either direction, but the concentration of solutes on either side of the membrane does not change.
When solutes are uncharged, each type of solute (e.g., glucose, oxygen) tends to move down its own concentration gradient independent of any other concentration gradient (Figure 5.18). A cell does not need to expend energy to transport solutes down (with) their concentration gradients, but it must use energy to move solutes up (against) their concentration gradients, as will be further discussion in Chapters 5.3 and 5.4.
Electrochemical Gradients (charged solutes)
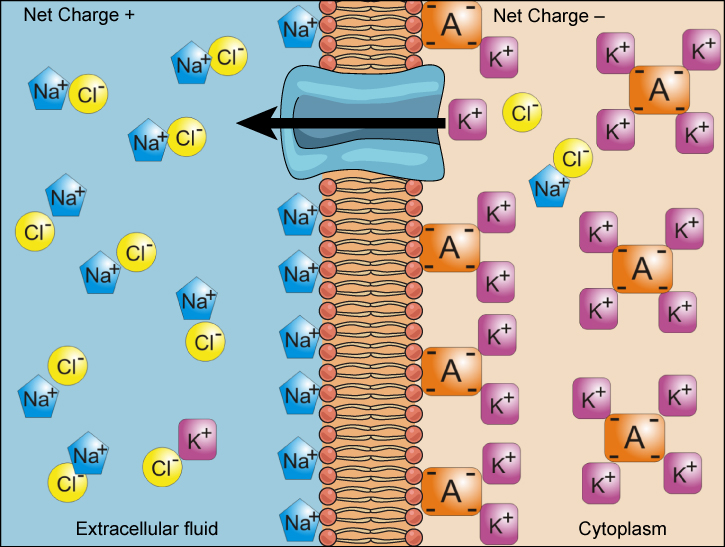
Some solutes are ions with positive or negative charges. These solutes contribute to an electrochemical gradient that exists across most cell membranes. An electrochemical gradient is an electrical gradient caused by an asymmetrical distribution of charged chemicals (i.e., solutes). This gradient depends not only on the concentration of solutes, but also on their charge. For example, positively-charged ions like sodium (Na+) increase the positive charge in an area, while negatively-charged ions like chloride (Cl–) do the opposite (increase the negative charge or describe the positive charge in an area). In an electrochemical gradient, one side of the membrane is more negatively charged, while the other is more positively charged. This difference in charges across the membrane is called a membrane potential (Vm).
A key difference between concentration gradients and electrochemical gradients is that charged solutes don’t move totally independently of other solute gradients. This is because charged solutes are attracted to oppositely charged regions. For example, the presence of lots of Cl– ions on one side of the membrane can influence the movement of Na+ ions, since they have opposite charges, and opposite charges attract each other. Thus we talk about electrochemical gradients rather than concentration gradients for ions because their movement is affected by their own chemical gradients, but also the gradients of other charged solutes.
We will delve further into this concept by analyzing the complexity of ions in a living system. The membranes of most cells have a negative membrane potential, suggesting an excess of negatively charged solutes in the cytosol, relative to the extracellular fluid. Most animal cells also have higher concentrations of potassium (K+) and lower concentrations of sodium (Na+) in the cytosol than in the extracellular fluid. If the appropriate channels exist in the cell membrane, Na+ tends to diffuse into the cell, down both a concentration gradient and an electrical gradient, toward the negatively charged cytosolic side of the membrane. The situation is more complex for other elements such as potassium. The concentration gradient of K+ drives K+ out of the cell, but it seems like the electrical gradient should drive K+ into the cell. However, in Figure 5.19 we see K+ is leaving a cell (towards the positively-charged extracellular fluid) because of the combination of concentration and electrical gradients.
To fully understand the example above, we must talk about equilibrium potential. Each ion has an equilibrium potential: a membrane potential at which there is no net movement of that ion across the membrane. This is a similar concept to that of dynamic equilibrium for neutral (uncharged) solutes. However, at the equilibrium potential, ion concentrations on either side of a membrane are usually still unequal, but electrical charges (membrane potential) prevent further net ion movement. For example, in a human neuron, the equilibrium potential for K+ is -88 mV, which can occur while there is still much more K+ inside the cell compared to outside the cell. As long as the current membrane potential is different from an ion’s equilibrium potential is different, that ion will tend to cross the membrane via appropriate protein channels. If, for example, the membrane potential of a cell is -70 mV, K+ will leave the cell (via K+ channels) down its electrochemical gradient until the membrane potential becomes -88 mV, at which point there should be no net movement of K+ across the membrane. Different ions have different equilibrium potentials. In the same neuron, Na+ has an equilibrium potential of +60 mV, and Cl– has an equilibrium potential of -61 mV.
Electrochemical gradients play a crucial role in the movement of specific ions for various operations such as neuron function and muscle contraction in animal cells. As a result, cells must invest energy into maintaining these electrochemical gradients. Electrochemical gradients are also important across some organelle membranes. For example, both mitochondria and chloroplasts maintain an electrochemical gradient driven by hydrogen ions (H+, protons) across the inner mitochondrial membrane and the thylakoid membranes, respectively.
