10.3 Strategies and Mechanisms of Oxidative Stress Tolerance
KEY CONCEPTS
By the end of this section, you will be able to do the following:
- Explain how the main strategies cells can use to combat oxidative stress are effective at minimizing the negative effects of ROS
- Give examples of two mechanisms that can be used to modify the balance between ROS production and neutralization
- Give examples of mechanisms that can be used to repair ROS damage to proteins, membrane lipids, and DNA.
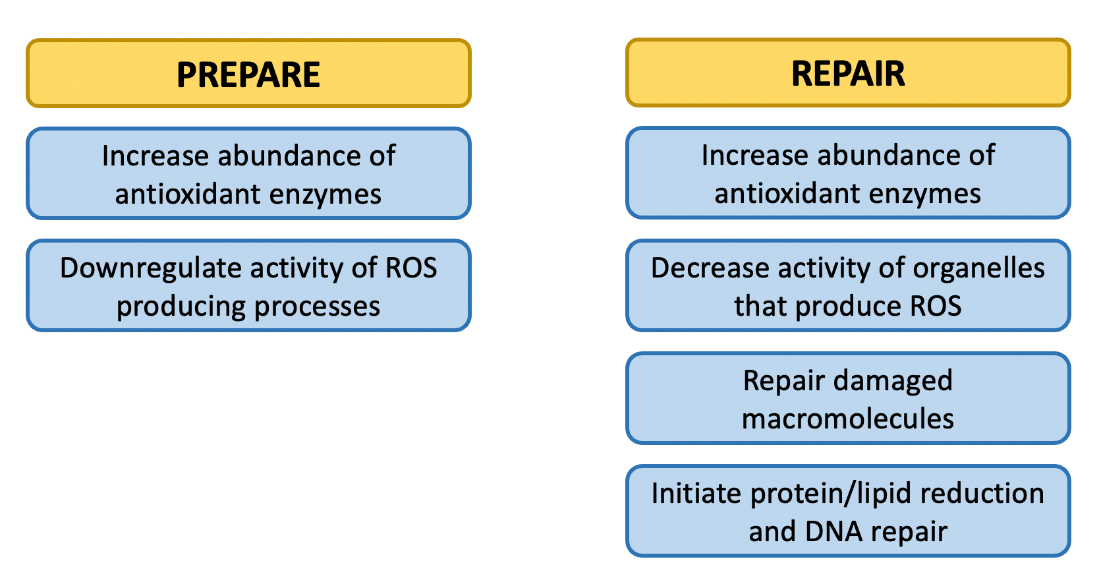
Once cells have detected oxidative stress, they can respond in several ways. Because oxidative stress is defined as an imbalance between ROS production and neutralization, we can organize our understanding of oxidative stress into two strategies: decrease ROS production or increase ROS neutralization. Both of these strategies can be enacted before and after an oxidative stress event (Figure 10.6). In addition, if ROS themselves cannot be controlled by the organism, a third strategy of repairing damage to macromolecules may be required. For example, if oxidative stress is a predictable part of an organism’s life cycle (e.g., a seasonal event), the organism may downregulate ROS production and increase antioxidant defenses prior to the predictable stress event. Alternatively, if the stress is unpredictable (e.g., an unexpected stroke), these strategies may only be enacted after the stress occurs. This latter option is usually more energetically costly than preparing for the stress in advance. In this chapter section, we will examine some of the mechanisms organisms can use to support the strategies of oxidative stress tolerance.
Decreasing ROS Production
In anticipation of stressful events, many organisms pre-emptively modify their metabolic pathways. For example, many animals enter a state of diapause prior to winter or other stressful conditions. Diapause is a type of developmental dormancy associated with very low metabolic rates and other adaptations for stress tolerance. The decrease in metabolic rates is often driven by suppression of mitochondrial activity, for example by shutting down the mitochondrial electron transport systems (ETS) or degrading mitochondria. [1]In the annual killifish (Austrofundulus limnaeus), the changes to mitochondrial ETS activity likely help suppress ROS production during and after stressful conditions.
Increasing Abundance/Activity of Antioxidant Enzymes
An increase in the abundance or activity of antioxidant enzymes can help confer oxidative stress either pre-emptively or as a response to an oxidation event. [2]For example, the freeze tolerant goldenrod gall fly, Eurosta solidaginis, overwinters at very low temperatures, so it’s body fluids freeze, and thawing can cause an increased in ROS production (as discussed in Chapter 10.1). The goldenrod gall fly is able to increase the activity of antioxidant enzymes, such as superoxide dismutase (SOD) and catalase (CAT) throughout the winter, so it is prepared for oxidative stress in the spring thaw (Figure 10.7).

Repairing Oxidized Macromolecules
When a cell is unable to predict the oxidative stress, they are unable to pre-emptively upregulate antioxidant enzymes, and must use other mechanisms to repair oxidized macromolecules. The cell can use different antioxidant enzymes to repair damaged macromolecules after an oxidative event. These families of antioxidant enzymes are able to reduce lipids and proteins that have been oxidized. Other mechanisms such as, base excision repair, are used to repair damaged DNA.
One example of protein repair in mammalian cells is through the use of glutaredoxin (Grx) and thioredoxin (Trx) enzymes. Grx and Trx are antioxidant enzyme families that catalyze various reactions to reduce oxidized proteins, thereby restoring proteins that have been reversibly oxidized by ROS. These enzymes are found in the cytosol, the mitochondria, the nucleus, and they can be secreted extracellularly. For reduction to occur, the active sites of Grx and Trx are oxidized while the target protein’s disulfide bonds (e.g., in cysteine, methionine) are reduced (Figure 10.8A, B). Then, Grx and Trx must be oxidized back to their original form, because enzymes should not be consumed by a reaction (Figure 10.8A, B). Repairing damaged proteins is only possible for mild oxidative stress that can be repaired quickly. As explained in Chapter 10.2, not all protein oxidation is reversible. Whenever proteins are irreparably damaged, they are typically degraded and replaced with newly synthesized proteins. The response to this type of damage will be covered in Chapter 14.
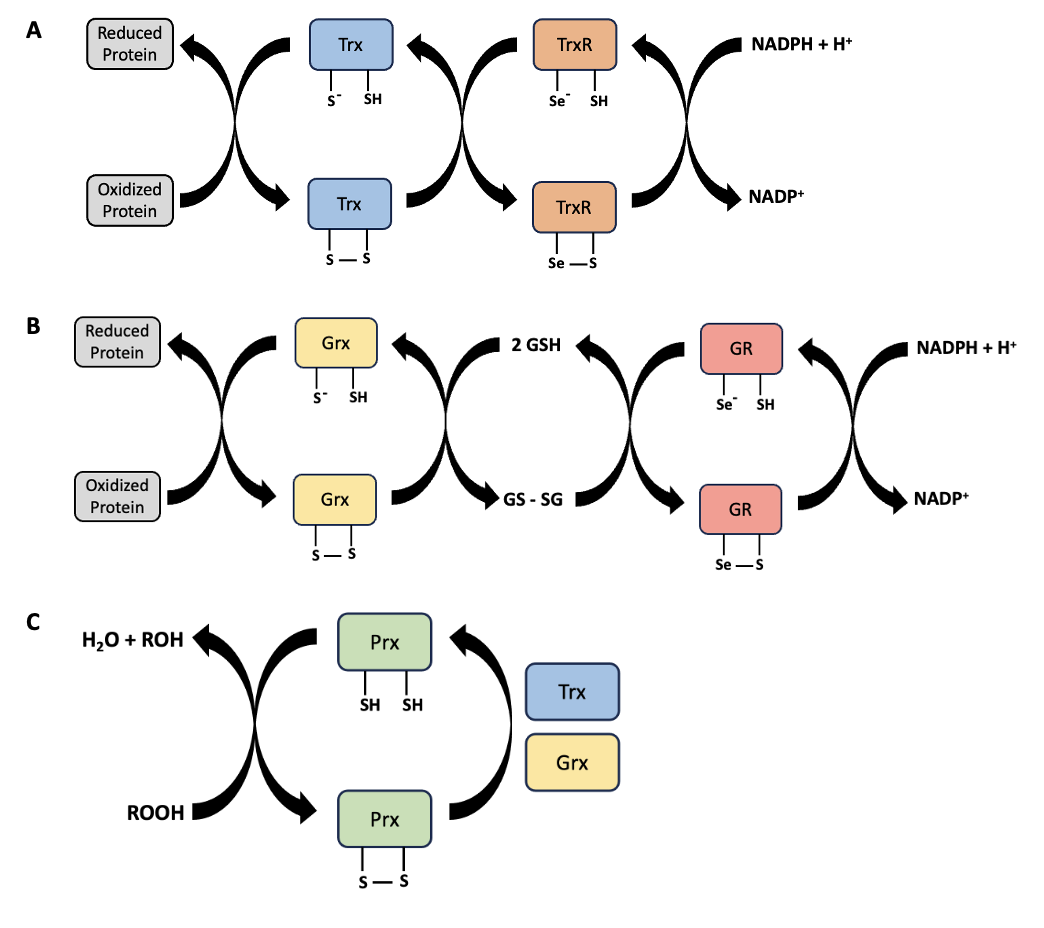
Some membrane lipid peroxidation can also be repaired by antioxidant enzymes. Peroxiredoxin (Prx) enzymes can catalyze the reduction of hydrogen peroxide to water, and can help neutralize lipid peroxides (Figure 10.8C). Similar to Grx and Trx, Prx itself gets oxidized when catalyzing the reduction of these peroxides. Prx is then an oxidized protein, and can be reduced to its normal state by enzymes like Grx and Trx (Figure 10.8A, B). Similar to repair of oxidized proteins, repair of lipid peroxides is time-sensitive and not all lipid peroxidation is reversible. Damaged membrane lipids may instead have to be replaced.
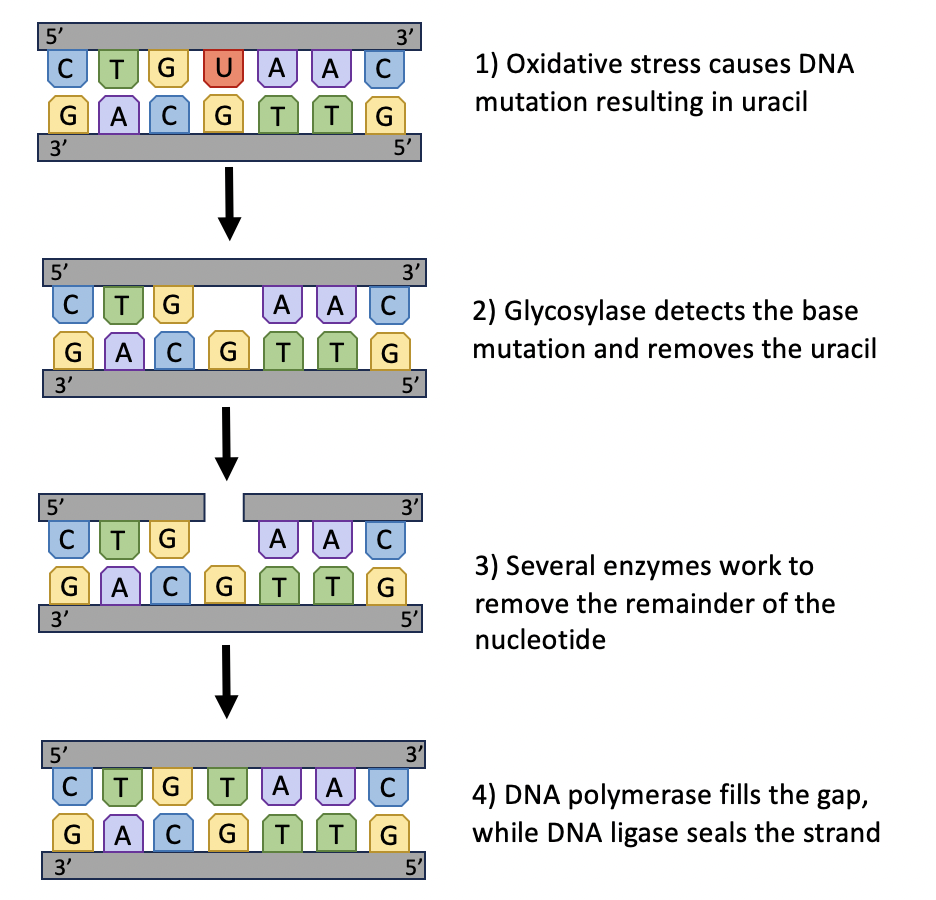
To repair oxidative damage to DNA, Base excision repair (BER) is a common repair pathway. BER can be used to combat single nucleotide mutations and other mild damage to DNA (Figure 10.9). The first step of involves detecting and removing the damaged bases; the enzyme glycosylase facilitates this process. Once glycosylase removes the base, several other enzymes remove the rest of the nucleotide from the DNA strand. Finally, the gap in the DNA backbone is filled and sealed with the help of DNA polymerase and ligase. This mechanism of repair can help restore DNA to its original form as long as more severe damage (e.g., breaks in the sugar-phosphate backbone) have not yet occurred.
- Duerr, J.M., and Podrabsky, J.E. 2010. Mitochondrial physiology of diapausing and developing embryos of the annual killifish Austrofundulus limnaeus: implications for extreme anoxia tolerance. J Comp Physiol B 180(7): 991–1003. doi:10.1007/s00360-010-0478-6. ↵
- Joanisse, D.R., and Storey, K.B. 1996. Oxidative Stress and Antioxidants in Overwintering Larvae of Cold-Hardy Goldenrod Gall Insects. Journal of Experimental Biology 199(7): 1483–1491. doi:10.1242/jeb.199.7.1483. ↵
