9 Chapter 9: Sensation & Perception – The Chemical Senses
COVER IMAGE
Ben Marcus, PhD (University of Chicago)
Austin Lim, PhD (DePaul University)
In the previous two chapters, we examined some of the ways we detect and perceive our physical environment, including through phenomena such as light, sound, and tactile sensations.
In this chapter, we will complete our tour of the senses by exploring how we sense and perceive chemical compounds. In particular, we will describe the neural processes underlying our senses of smell, taste, and the chemical status of our internal homeostatic state.
Our olfactory system (smell) is activated by compounds called odorants, while our gustatory system (taste) is activated by compounds called flavorants. These two chemical senses are closely intertwined; odorants affect how we perceive taste, and bad tasting foods often smell bad. Try eating a fancy meal with a stuffed-up nose, and you’ll quickly notice that you lose the subtle complexities (and most of the joy!) of those foods.
Chemicals in our body, such as the amount of carbon dioxide in the blood or the presence of toxins in our digestive tract, are detected with our internal chemosensory systems. Many of these sensory systems are intimately tied with our autonomic nervous system (Chapter 2). Changes in chemical balance here results in unconscious or involuntary physiological changes to restore homeostasis.
Chapter 9 outline
9.1 Olfactory System
9.2 Gustatory System
9.3 Internal Chemosensory Systems
9.1 Olfactory System
Olfaction is the ability to sense and perceive volatile chemicals that are suspended in the air. The typical human can distinguish up to 10,000 distinct odors, ranging from the sweet aroma of esters produced by apples and oranges, to the putrid smells of sulfurous compounds produced by skunks and rotten eggs. Because odors can drift along in the air, some chemicals can be detected long before the source is within eyesight: think about smelling a burning bonfire from miles away.
Smells affect our conscious behavior. They can motivate us to approach freshly-baked bread or avoid a rotting animal carcass. These chemicals serve as survival cues: bread gives us energy-rich carbohydrates while a decaying carcass can expose us to disease. Odorants can even affect our behaviors unconsciously: sleeping amidst the scent of a romantic partner increases the efficiency of our sleep, and subliminal exposure to citrus smells can lead people to clean their space more completely after eating a crumbly biscuit (an effect described in a study called “Smells Like Clean Spirit,” a nod to 90s punk band Nirvana).

Olfaction is one of the oldest functions we possess as animals. While our sense of smell has seemed to take a backseat to other senses relative to other animals (about 2% of the total mouse brain mediates smell, while only a scant 0.01% of the volume of the human brain is dedicated to this function), scientists now appreciate that our olfactory system is simply more specialized. For example, humans use the smells of sweat to clue us into the emotional state of others, and we can even subconsciously detect sickness through body odor.
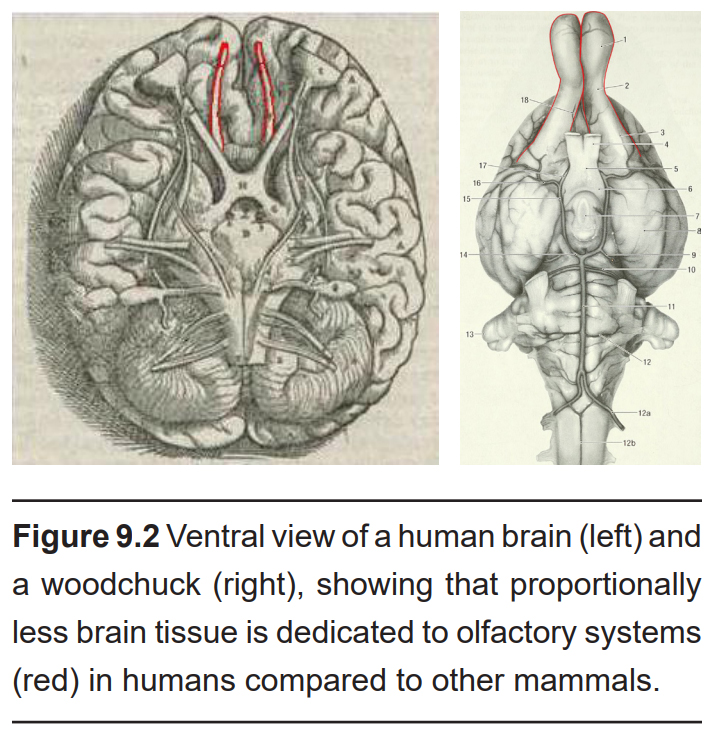
9.1.1. Anatomy of the Olfactory System
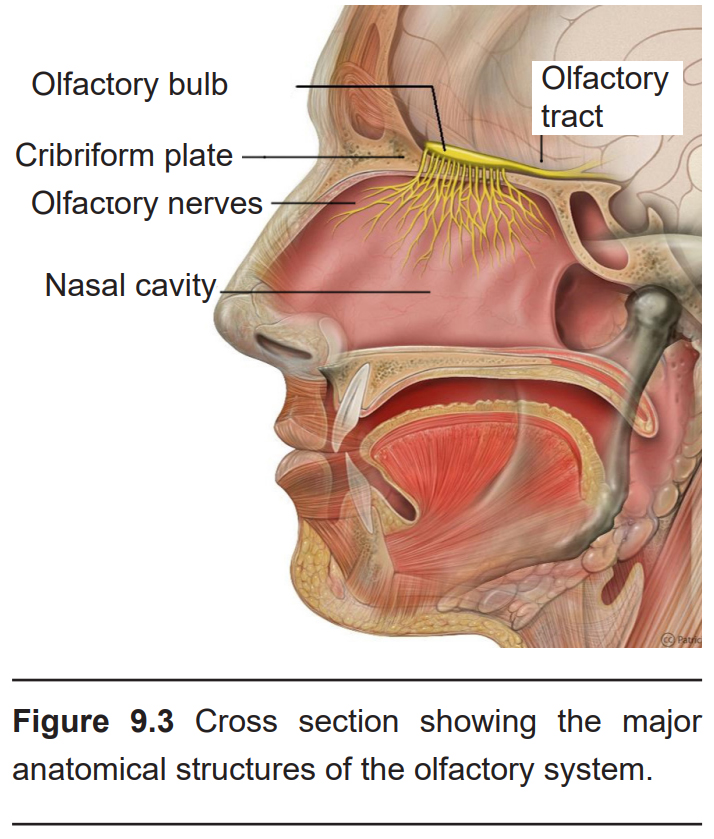
Your sensation of smell begins when an odorant traverses your nostrils and passes through the nasal cavity, an empty, air-filled space just behind the front of the skull. The dorsal-most portion of the nasal cavity is covered in a mucus-covered patch of tissue called the olfactory epithelium.
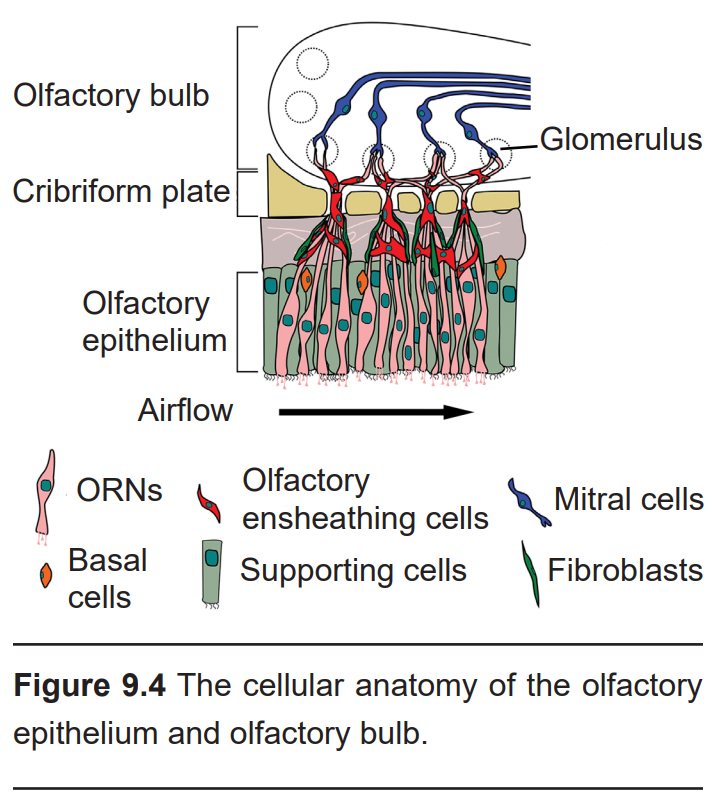
Embedded within the olfactory epithelium are the olfactory receptor neurons (ORNs, also called olfactory sensory neurons, or OSNs), neurons which begin processing smell. ORNs are bipolar neurons, with long dendritic projections protruding into the epithelium. Here, they peek out and contact the air, making them the only neurons that are directly exposed to the outside world. This, unfortunately, causes them to encounter all sorts of dangers such as toxins, particulates, and microbes. They are one of the few known populations of neurons where adult neurogenesis occurs, each having a lifespan in the range of 30 days to a year. The average human olfactory system has somewhere between 6-20 million ORNs.
Within the olfactory epithelium are a population of supporting cells, which function much like glial cells. They help dispose of dead and dying cells, metabolize pollutants, and may also help to physically maintain the epithelium.
On the dendrites of ORNs are olfactory receptors. It is estimated there are about 1,000 different genes (about 3% of the total human genome) that code for roughly 400 different olfactory receptors. Each ORN expresses only one type of olfactory receptor, and each receptor is believed to respond to activation by a different chemical. For example, the receptors expressed on ORN293s are highly responsive to cadaverine, the smell characteristic of death and decay. Even though scientists have identified the odorants that activate some receptors, most of the olfactory receptors are still “orphaned,” and have not yet been matched with their corresponding odorant. The initial research into the genetics underlying these neurons earned Drs. Linda Buck and Richard Axel a Nobel prize in 2004, but there is still much work to be done.
Olfactory receptors are transmembrane G protein-coupled receptors that signal downstream effectors using the intracellular transduction molecule Gαolf. This protein complex is 90% similar to the stimulatory G-protein Gs, and likewise triggers activation of adenylate cyclase, elevating the intracellular concentration of cyclic AMP (see section 5.3 for a refresher on this signaling pathway). Activation of Gαolf causes depolarization, causing the ORN to fire action potentials.
ORNs encode the intensity of smells through the frequency of action potential firing, which changes in accordance with the concentration of odorant molecules. Imagine standing over a fresh-baked pizza and inhaling deeply. Due to the high concentration of odorant molecules in the air, several receptors will be activated, leading to frequent neuronal firing. Now, imagine you are down the block from a pizza restaurant, getting only a slight whiff of those same scents. Here, the concentration of odorants is low, meaning that the ORNs fire less frequently.
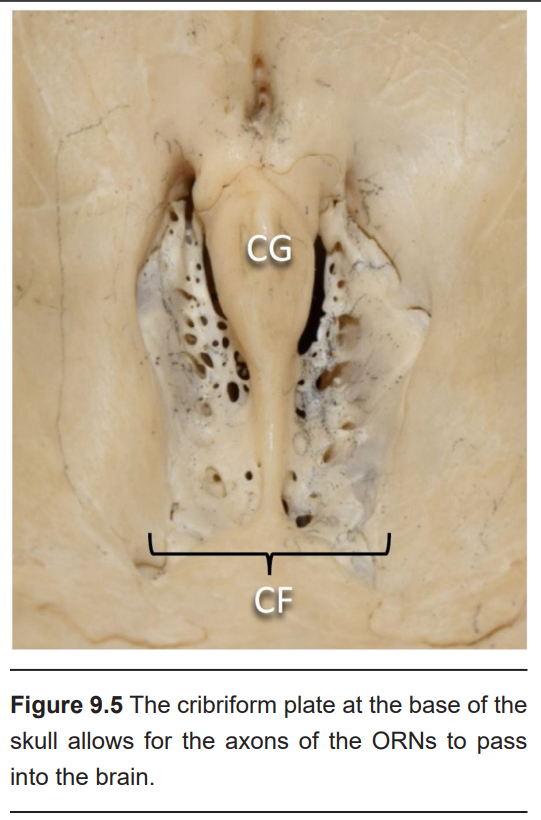
Following activation of the receptors, the axons of the ORNs pass through the skull through a tiny series of holes at the cribriform plate, a sieve-like section of the ethmoid bone. These primary neurons form synaptic connections onto neurons in the olfactory bulb, the beginning of the olfactory nerve (CN I). Like the optic nerve, the olfactory nerve runs along the ventral surface of the brain.
The site of synaptic connectivity between the ORNs and the secondary neurons in the olfactory bulb is a highly specialized clump of tissue called a glomerulus. The typical human has a little under 2,000 glomeruli, and each glomerulus only receives inputs from ORNs that express the same type of olfactory receptors.
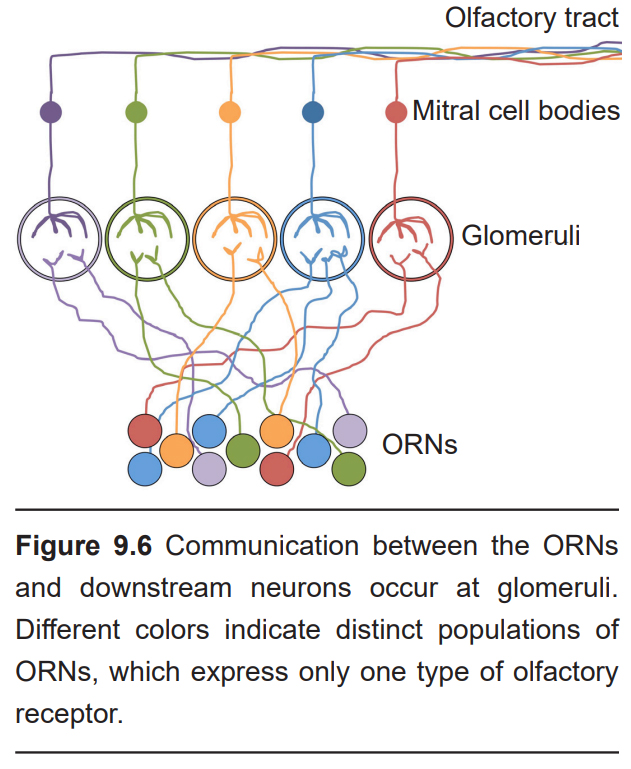
Within each glomerulus are the dendrites of the secondary neurons, which exist in two types: The mitral cells and the tufted cells. These two cell populations project axons directly into the olfactory cortex. This makes the olfactory system the only sensory system that does not pass signals through the thalamus before cortical processing.
There are also two types of inhibitory neurons that regulate this pathway: granule cells found within glomeruli, and periglomerular cells, which send axonal projections into the glomeruli, both help refine the synaptic processing of scent information using lateral inhibition, a system similar to that found in the retina of the visual system (Chapter 7).
Collectively, the olfactory cortex is made up of several different regions.
- Piriform cortex
The piriform cortex is the main cortical input site for axonal projections from the olfactory bulb. In some species of mammals that rely very heavily on their sense of smell for survival, up to 10% of total cortical volume is piriform cortex. The outputs of the piriform cortex project into other structures of the olfactory cortex, as well as the mediodorsal thalamus, a relay center that contributes to learning and decision making.
- Cortical amygdala
The amygdala is a part of the brain that helps mediate complex emotional states (chapter 15). Generally, it is subdivided into a few different subnuclei. One of them in particular, the cortical amygdala, receives strong inputs from the olfactory nerve. The cortical amygdala sends projections into the hippocampus, a brain structure critically important for the formation of new memories.
The strong neural connections between the olfactory nerve and amygdala are believed to be the reason why smell and memory are strongly linked. Think about a time when you caught a slightest whiff of a scent, and that scent mentally transported you to a distant place from a long time ago (this experience was also described by author Marcel Proust in his highly- regarded novel, In Search of Lost Time.) We form associations between specific smells and emotionally salient memories. Some odorants, like the volatile Maillard reaction products that are made as a product of cooking, remind us of the “good ole days” watching a parent bake or grill. Many bakeries take advantage of these positive memories and point their exhaust out into the street to entice passersby.

Unfortunately, the memories associated with smells can also have negative valence. For example, the smell of petroleum and oil fumes can trigger a sickness-like conditioned response in Gulf War veterans.
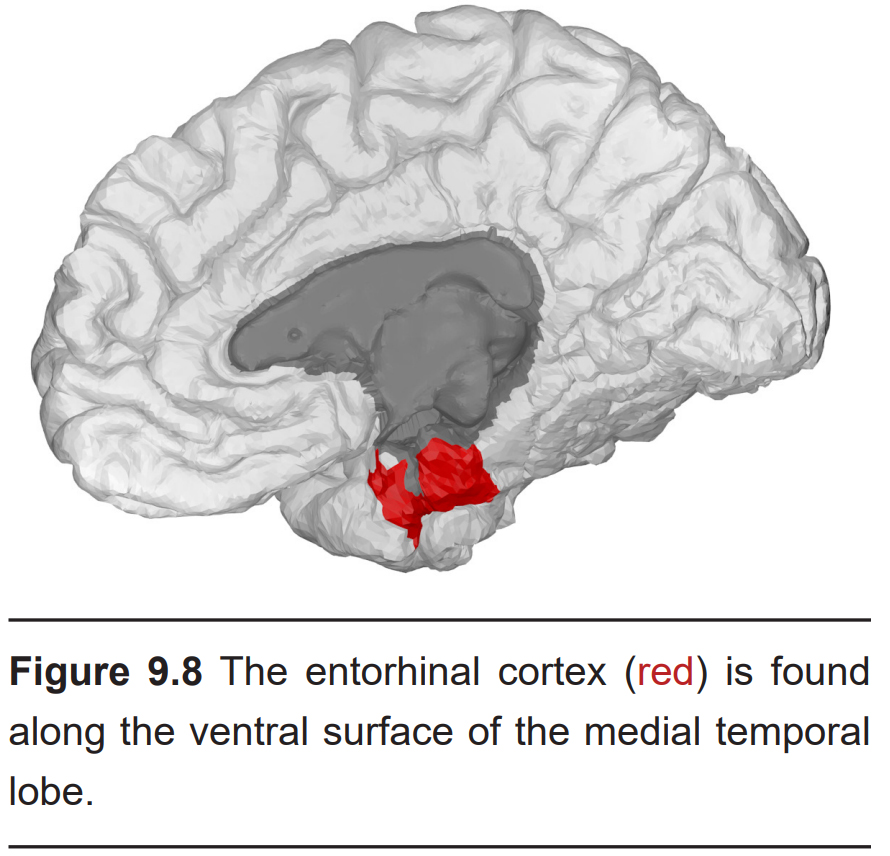
- Entorhinal cortex (EC)
The entorhinal cortex is a small section of the medial temporal lobe. As with the cortical amygdala, the EC sends strong connections into the hippocampus, indicating that olfactory signals contribute to the strong associations formed between smell and memory through EC, as well.
Other than receiving and processing olfactory signals, the entorhinal cortex is also involved in spatial navigational tasks.
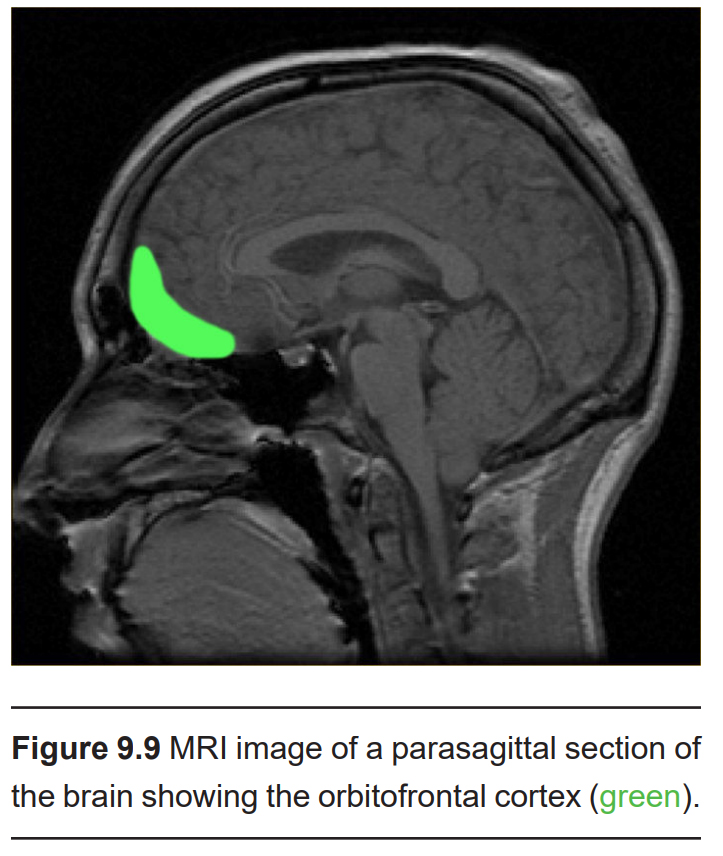
- Orbitofrontal cortex (OFC)
As the name implies, the orbitofrontal cortex (OFC) is found just behind the orbit, the bony socket of the skull where the eyes sit. OFC is found at the ventral surface of the frontal lobe. Circuits in this brain area function as an integration site for sensory inputs, since it also receives projections from visual, taste, and somatosensory cortices.
The full extent of the OFC is still under examination, but it is also implicated in decision making and social behaviors.
Do humans respond to pheromones?
Some species secrete chemicals which influence the behavior of other members of the same species. These pheromones trigger complex responses in others, such as motivating social insects like ants to follow meandering trails towards faraway food sources or causing a group of bees to swarm and attack a predator.
There is still debate over whether or not humans produce and release pheromones, and whether or not other humans are sensitive to these signals. Many nonhumans, notably snakes and dogs, have a specialized structure in their nasal cavity called the vomeronasal organ, which detects organic compounds produced by predators and reproduction- related hormones produced by the opposite sex. These signals are sent into the brain via cranial nerve 0, which can then trigger behavioral changes. Humans have these anatomical structures, but they are not believed to carry any functional information. However, other evidence suggests that smelling certain compounds influences mood, alters neuroendocrine signaling, and even affects mate selection.
9.1.2 Disorders of the Olfactory System
Like other sensory systems, the structures involved in olfaction can be injured. An injury to the olfactory system can result in hyposmia, a reduced ability to smell, or anosmia, a complete loss of smell.
The most common insult to the olfactory system is simple nasal congestion, a temporary, physical blockage of the entry to the nasal cavity that decreases airflow and, therefore, the number of particles that reach the olfactory epithelium. Congestion can be caused by allergies, the common cold, upper respiratory bacterial or viral infections, or sinus infections. Hyposmia is also one of the main neurological symptoms of COVID-19.
Hyposmia is common among healthy, older adults, affecting about half of the population between 65 and 80 years old. As a person ages, spontaneous calcification causes the holes in the cribriform plate to shrink, which can impinge on and damage ORN axons.
Hyposmia can also be caused by abrupt head injuries. The ORN axons that project through the holes of the cribriform plate are particularly sensitive to blows to the head.
Neurodegenerative disorders, such as Parkinson’s disease and Alzheimer’s disease, contribute to smell deficiency. Usually, hyposmia precedes the major clinically observed symptoms of these disorders, hinting that smell deficiency may serve as an early diagnostic biomarker.
Another olfactory deficit, phantosmia, is when a person perceives “phantom” scents, or in other words, experiences an olfactory hallucination. Phantosmia may be triggered by a temporal lobe seizure or a stroke. It can also be caused by a brain tumor affecting the olfactory nerve (CN I), or the subsequent surgical removal of the tumor, leading to injury. Schizophrenia, a psychiatric condition characterized by auditory hallucinations, may also cause phantosmia.
9.2 Gustatory System
Your gustatory system, which mediates your sense of taste, helps you walk the line between health and illness. It guides you towards foods that are energy rich, and keeps you away from food that could make you sick. The specific taste modalities, salt, sweet, sour, bitter, and umami, all support this balance. Sweet foods taste good because foods rich in sugar, such as fruit and wheat, contain large amounts of usable energy, and humans have evolved to find these foods appetizing. In contrast, toxic compounds are often bitter, causing you to respond with feelings of disgust.
To further refine this balance, a tastant’s appeal varies depending on its concentration. For example, we find a small amount of sourness desirable in fruit and candy, but aversive in milk. And while bitterness is often a sign of toxicity in food, mild bitterness contributes positively to the taste of coffee and chocolate. Sugar can make candy, pastries, and drinks very appealing, but too much can make the food barely palatable. The same applies to salt: the right amount makes potato chips and pretzels delicious, but it also causes sea water to make you vomit. All these reactions are adaptations humans have developed to maintain homeostasis in our body.
9.2.1 Anatomy of the gustatory system
Lingual papillae (singular; papilla) are the large anatomical structures that give the tongue its characteristic rough surface. These structures can be seen with the unaided eye. Each papilla contains up to a hundred taste buds, which are onion-shaped taste receptors. Taste buds can also be found on the palate and in the throat. In total, a person has about 10,000 of these receptors, but the number varies by age: Taste bud concentration in the mouth peaks in childhood and decreases throughout adulthood.
Within each taste bud are approximately 100 taste receptor cells. Technically, these cells are not neurons, but are instead derived from specialized epithelial cells. These cells reach toward the apical tip of the taste bud and sprout thin projections called taste hairs, which extend into taste pores. These tiny pockets at the apical tip of the taste bud are where taste hairs meet saliva. Taste buds also contain basal cells, which reproduce to form supporting cells, and over time, these mature further into taste receptor cells. Taste receptor cells turnover every 8–22 days.
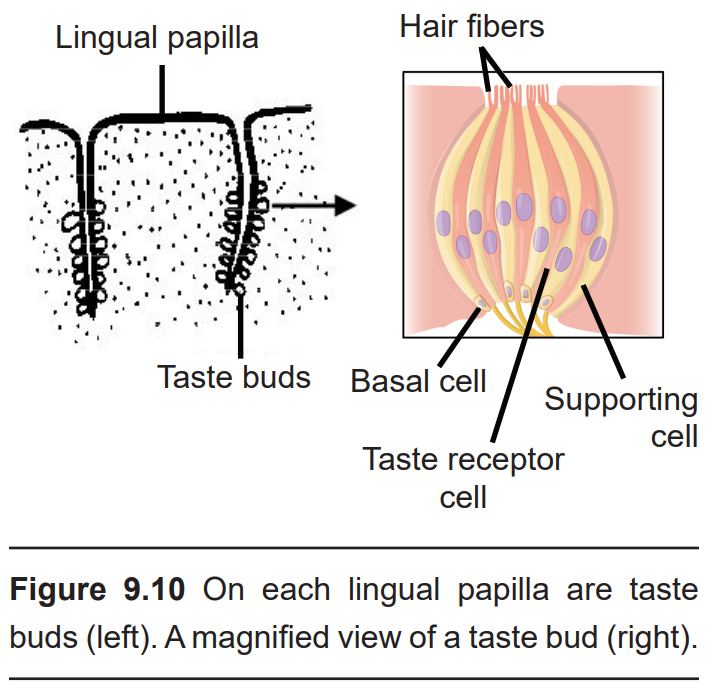
Taste receptors cells are responsible for sensing and conveying information about taste in accordance with the main taste modalities (salty, sweet, sour, etc). Like how the olfactory system is organized, taste receptor cells do not express receptors for more than one taste modality – a salty taste receptor cell only carries salty information, a sweet taste receptor cell only pass on sweet information, and so on.
Taste receptor cells communicate with afferent gustatory neurons. These gustatory nerve fibers originate from three of the twelve cranial nerves.
- The nerve fibers from the anterior two- thirds of the tongue are part of the facial nerve (CN VII).
- The posterior third of tongue sends information through the glossopharyngeal nerve (CN IX).
- The back of the palate and the throat can send taste-related signals through the vagus nerve (CN X).
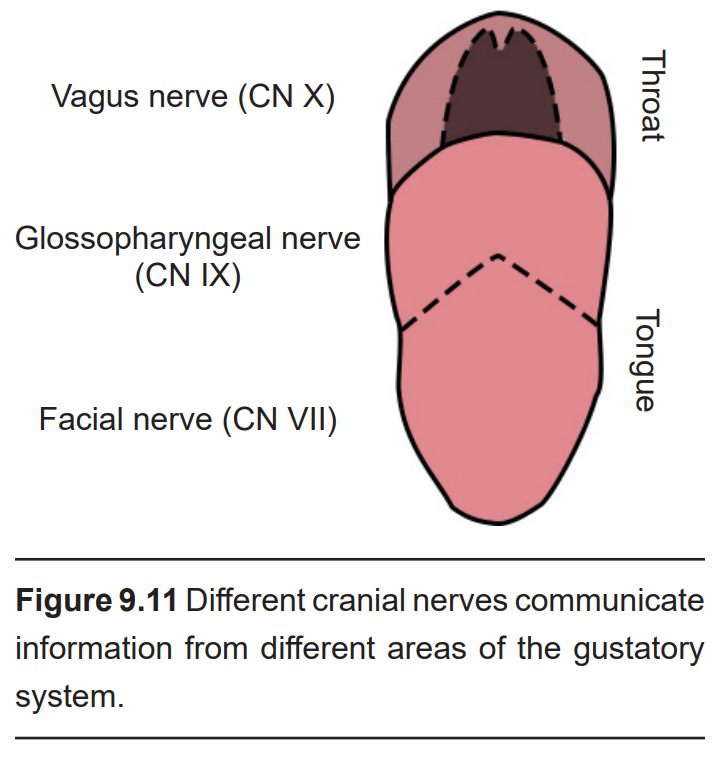
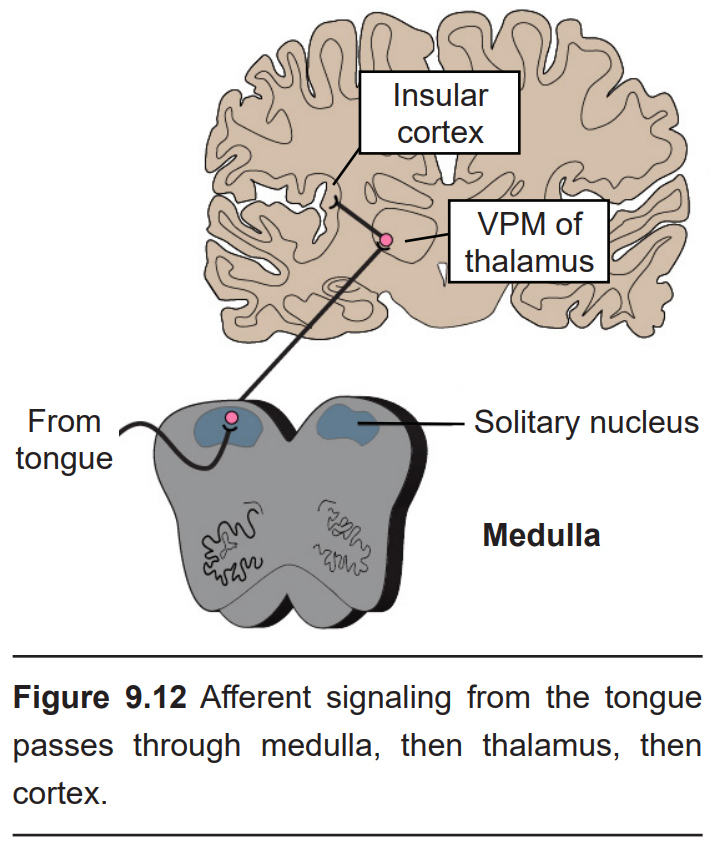
These neurons then form synapses on second order neurons in the rostral medulla in an area called the solitary nucleus (or gustatory nucleus) in the medulla oblongata. Unlike most other sensorimotor systems, the gustatory system sends ipsilateral projections into the CNS; that is to say, taste information from the left half of the tongue gets represented in the left hemisphere of the brain, and visa versa.
From the medulla, these neurons send axonal projections into the ventral posteromedial (VPM) nucleus of the thalamus. (These represent the third-order neurons of the gustatory system.) These neurons send projections widely across several areas of the cortex.
The gustatory cortex is the beginning of the processing of taste perception. It is made up of two different parts: the anterior end of the insular cortex and the frontal operculum of the frontal lobe. These neurons convey information such as the taste modality and intensity.
9.2.2 Taste modalities of the gustatory system
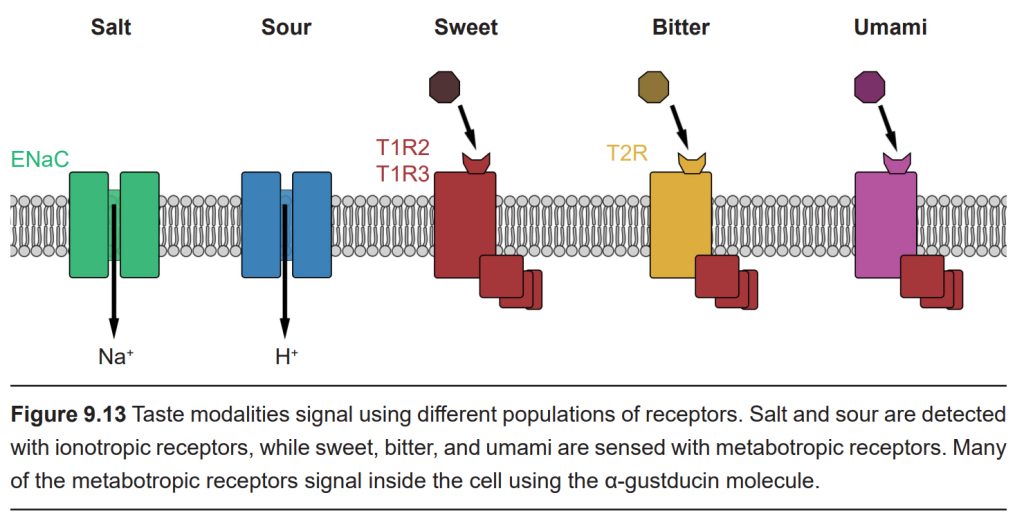
Currently, it is believed that humans sense five basic tastes: Salty, sour, sweet, bitter, and umami. Salty and sour taste are both mediated by ionotropic taste receptors, while sweet, bitter, and umami taste are mediated by metabotropic taste receptors. Besides varying by type, these receptors are also not distributed evenly across the surface of the tongue. For example, the tip of the tongue is most sensitive to sugars, while the back of the tongue is most sensitive to bitter compounds.
Salt
Sensation of salt taste is primarily driven by Na+ ions. When you sprinkle a little table salt (NaCl) onto your tongue, it dissolves in saliva and the free sodium ions can passively influx into salt taste receptor cells through epithelial sodium channels (ENaCs). This movement of positively charged Na+ ions causes depolarization of the taste receptor cell, just like in neurons. This depolarization activates voltage-gated calcium channels, prompting neurotransmitter release that, in turn, activates the afferent gustatory nerve fibers.
Salty foods elicit a biphasic response depending on concentration. Foods cooked with a low concentration of salt taste bland and are not very appetizing; however, high salt concentrations elicit a strong aversive reaction – imagine how disgusted you were when you first tried to cook and were overly generous with the salt! The taste of salt is typically desirable at salt concentrations lower than 100 mM.
Additionally, the appeal of salt at a given moment depends on our body’s need for salt at the time. Several hormones such as the appetite-stimulating hormone ghrelin contribute to regulating the concentration of salt in the body by mediating Na+ absorption. Current salt levels can also impact appetite for salt. For example, in chronically-sodium deprived animals, high salt solutions are highly rewarding.
Why are we so sensitive to the taste of salt? As it turns out, both Na+ and Cl− are essential nutrients. They are critical for maintaining blood volume and pressure, for regulating body water, for maintaining muscle contractions, and mediating action potentials. Cl-, in particular, helps maintain a healthy pH balance. But for these functions to be performed optimally, salt must be present at a specific range of concentrations in the body.
Sour
Sensation of sour taste is mediated by proton-selective channels, which is why acids are sour (acids are low pH, which means a high concentration of H+). Like neurons, taste cells can depolarize with the entry of positively-charged ions, which influences signaling passed through the downstream nerve fibers.
The purpose for our ability to sense acids in our food is under debate. Sour tastants do not inherently provide any nutritional value, except in the case of Vitamin C. Humans and other higher primates cannot synthesize Vitamin C on their own, so it’s possible that we evolved to find a combination of sourness and sweetness attractive enough to consistently consume Vitamin C-rich fruits. However, sour can be aversive, motivating us to avoid spoiled or unripe foods that might contain pathogens.
Sweet
Sweet taste transduction is carried out by the activation of G-protein coupled receptors expressed on the sweet taste receptor cells. The most likely candidate for the primary sweet- sensitive GPCR is a heterodimeric receptor with the subunits taste receptor type 1 member 2 (TAS1R2 or T1R2) and taste receptor type 1 member 3 (TAS1R3 or T1R3). All sweet substances, including carbohydrates and artificial sweeteners, activate the taste pathway through these T1R2/T1R3 receptors. Interestingly, these receptors are also found in other parts of the body, including in the brain, pancreas, gastrointestinal tract, and fat tissue. While its function in these areas is not known, it’s theorized that it plays a role in glucose homeostasis.
Sugars like glucose and sucrose are essential for the survival of a species, since they are the main source of cellular energy. Therefore, our ability to detect sweetness plays a central role in regulating how much energy we take into our bodies. Of all the taste modalities, sweet is the strongest driver of food selection.
Bitter
Bitter taste is sensed via the T2R receptor. From here, the molecular signaling cascade branches off into two distinct pathways. The first pathway is similar to the sweet taste pathway; it recruits a similar set of signaling proteins en route to the afferent gustatory nerves, and knocking out any of the receptors along the sweet taste transduction pathway also attenuates bitter taste transduction. The second pathway uses a taste-specific phosphodiesterase, an enzyme that lowers the intracellular concentration of the signaling molecule cAMP. Some bitter compounds bypass the T2R receptor altogether, permeating the cell membrane and directly activating the GPCR. The bitter compound quinine, found in tonic water, does this. In high concentrations, quinine can cause disgust and vomiting.
Bitter taste prompts avoidance because bitter tastes often indicate toxicity. For example, cucurbitacins, a group of bitter-tasting compounds found naturally in gourds, including cucumbers and pumpkins, are believed to have evolved as an antiherbivory defense mechanism to prevent predation. The exact mechanisms that connect toxic compounds to disgust and vomiting are unknown; it’s possible that there is not a causative relationship among these reactions to toxins, but rather that humans evolved to respond to toxic compounds at multiple levels in parallel: in the mouth, through bitter taste, in the stomach with vomiting, and in the colon, with diarrhea.
Umami
Umami is the taste of savory deliciousness, such as the taste of rich chicken broth, a perfect medium-rare steak, or aged cheese. The word is derived from the Japanese word umai, which means “delicious.” Like sweet flavor, umami taste is mediated by a heterodimeric metabotropic receptor. Umami is signaled when a molecule of glutamate (chemically the same as the neurotransmitter!) binds to T1R1/T1R3 receptors.
The intracellular signaling transduction process is similar for sweet, bitter, and umami sensation. Tastants of these modalities all activate GPCRs that use the G protein α-gustducin. This activation increases the activity of phospholipase C-β2 (PLC-β2), which, in turn, activates the inositol 1,4,5-triphosphate (IP3) receptor, causing the release of calcium into the cytoplasm. The calcium opens the transient receptor potential cation channel subfamily M member 5 (TRPMP5), which causes the taste cell membrane to depolarize, generating an action potential. This causes the release of ATP into the synapse, which activates afferent nerve fibers to signal the presence of these tastants.
Through evolution, we have come to prefer the taste of umami because glutamate is a byproduct of cooking food. Cooking foods changes their chemical properties, thereby improving digestion, reducing toxicity, and increasing absorption of nutrients. Glutamate is one of the byproducts in the process of heating a food, and so it benefits us to appreciate these flavors.
Fat
Atriglyceride is a chemical made up of three fatty acids bonded to a glycerol molecule. They are a large part of our diet and are commonly found in animal fat and butter. Triglycerides contribute to the “mouthfeel” of foods, giving foods a creamy, rich quality. However, triglycerides themselves don’t have a taste.
In 2015, researchers found that humans might be able to sense fatty acid chain molecules in the mouth. It turns out that in their study participants could distinguish between a control drink, a bitter drink, a sour drink, and drinks containing fat. Participants initially grouped the fatty drinks with the bitter and sour tastes, suggesting that fat by itself is an aversive flavor, despite being high in energy content.
The research on this taste modality, called oleogustus, is still in its infancy compared to that of the other taste modalities. Researchers have not elucidated the mechanisms underlying this pathway.
Spicy
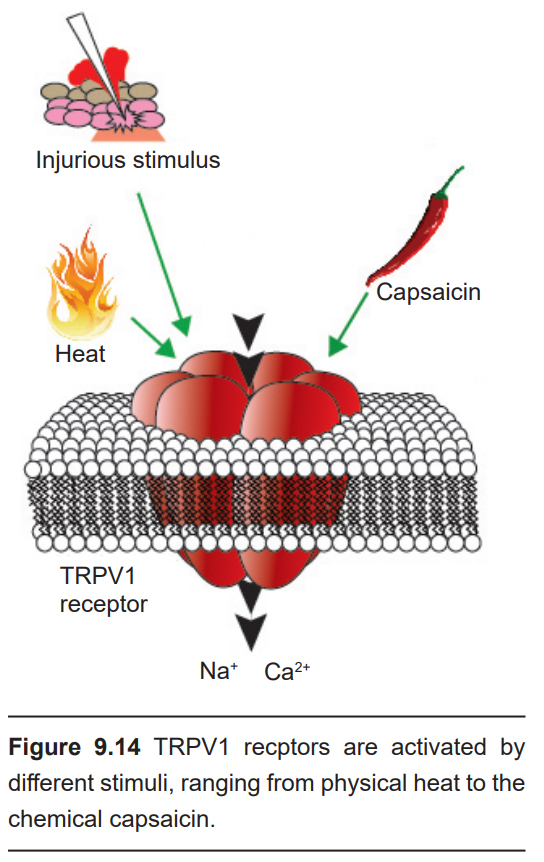
It’s no accident that the word “hot” is used to describe both the temperature and the spiciness of a food. After eating that delicious plate of curry, your body may exhibit a strong somatic response: intense salvation, a flushing of the skin, sweating, and sometimes even crying. As it turns out, the pain of eating a hot pepper is similar to experiencing other forms of physical injury. Taste receptor cells send signals to the brain via C fibers, the same neuronal pathway that carries afferent painful information (chapter 8).
Our ability to detect spicy flavors originates at the TRPV1 receptor, a nonselective cation channel. Upon activation, these receptors cause depolarization of the taste cell. Peppers contain the compound capsaicin, a potent activator of the TRPV1 receptor. These TRPV1 receptors are also temperature sensitive, opening at around 43C. Because these receptors can be activated by either chemical ligands or high temperatures, biting into a ghost pepper causes a similar sensation as if your tongue was literally being burned – the “heat” of spicy foods is more than just a colloquialism.
TRPV1 receptors are not only activated by capsaicin. Ethanol, for example, can also activate these receptors, which is why a shot of hard liquor causes a painful, burning sensation. Since the TRPV1 receptors permit proton movement into the cell upon activation, acidic conditions potentiate receptor activation, which also accounts for why acids often taste “hot.”
(Can we fix the spelling of receptor?)
9.3 Internal Chemosensory Systems
In addition to detecting chemicals with our noses or mouths, we also host a variety of chemosensory systems that sense various conditions about our internal environment. These systems contribute to the maintenance of homeostasis. Whenever the body is pushed out of it’s ideal operating range, these chemosensory systems respond by reflexively adjusting chemical absorption or behavior.
9.3.1 Respiration
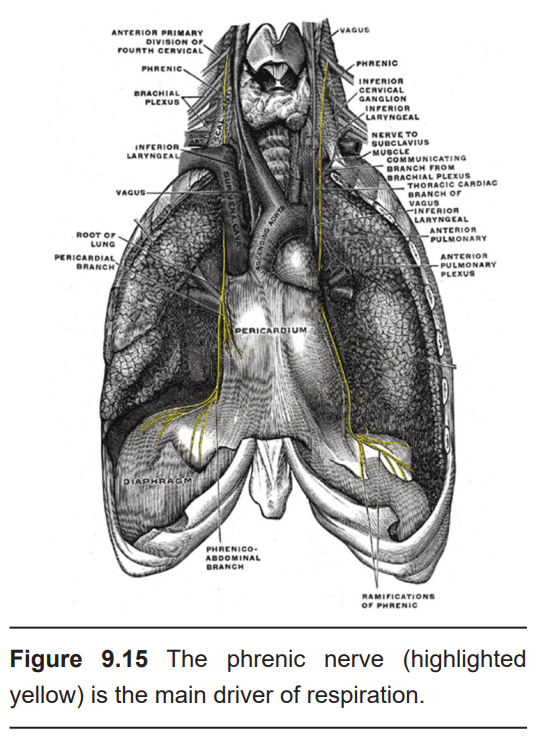
Neural control of the respiratory system originates in several circuits within the hindbrain. Of particular relevance is the medulla, the inferior- most segment of the brain stem. These complex circuits communicate with descending motor signals that are critically important for respiration through the action of two main nerves. The main driver of respiration is the phrenic nerve, which is the only nerve that innervates the diaphragm. The other drivers of respiration are the intercostal nerves, which innervate the intercostal muscles, the set of accessory respiratory muscles found between the ribs that expand the chest cavity during inhalation. People with spinal cord injury at the level of C5 or higher, which results in damage to the phrenic nerve, may need to be put on a ventilator. These circuits express opioid receptors, which is why opioid overdose can lead to fatal respiratory depression.
Regular respiration is an autonomic function. When CO2 levels rise (a condition called hypercapnia), these hindbrain neurons drive increased respiratory rate, which helps the body expel excess CO2.
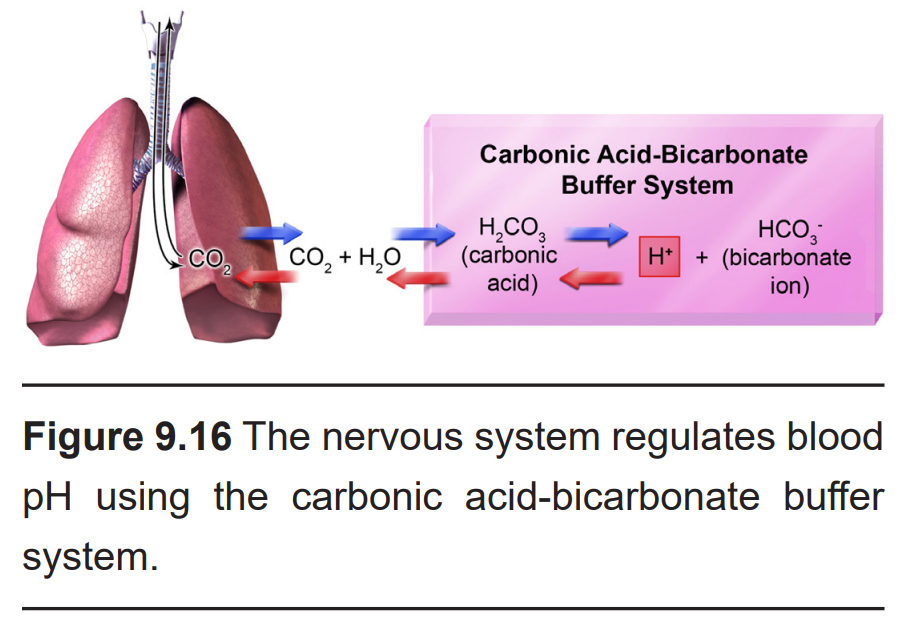
Respiratory patterns are also regulated homeostatically to restore a healthy level of pH in the blood. The pH of CSF is essentially a proxy measure for CO2 in the blood: CO2 diffuses easily across the blood brain barrier into the CSF. Once there, CO2 quickly reacts with H2O to form carbonic acid, which then dissociates into a bicarbonate ion and a H+ ion. Because of this chemical reaction, when blood CO2 is elevated, so is the concentration of H+ (low pH) in CSF.
Central chemoreceptors detect changes in the pH level of the CSF by sensing H+ ions, which enter the cells through acid-sensing ion channels (ASICs). When ASIC-expressing neurons detect pH levels less than 7, they send signals to the nerves that mediate diaphragm and intercostal muscle activity to increase respiration. This increases the exchange of CO2 out of the lungs, shifting the pH of the blood back towards more physiological levels of 7.4.
9.3.2. Vomiting
Vomiting (or emesis) is a rapid contraction of respiratory and abdominal muscles, compressing the stomach, thereby expelling stomach contents through the esophagus. Vomiting is often preceded by nausea, the unpleasant sensation of stomach discomfort.
Although aversive and painful, vomiting can be a natural and healthy protective response. For example, when toxins are produced during bacterial gastroenteritis (food poisoning), it is beneficial to expel the spoiled or rotten food from the stomach to minimize further exposure to bacterial toxins.
The neural signals that lead to vomiting originate at the afferent inputs of the vagus nerve (Cranial Nerve X), found in the intestinal tract. These ascending inputs form connections within the dorsal vagal complex (DVC), a series of nuclei found in the medulla of the brain stem. A region within the DVC that mediates the vomiting response is area postrema (AP), which is found on the floor of the fourth ventricle. Within AP is the chemosensory trigger zone, which is dense with neurons that sense the presence of various chemicals. The AP is considered to be a circumventricular organ, meaning that it is not isolated from the blood by a blood brain barrier. Instead, toxins and other large molecules in the blood are able to influence AP neurons directly. Additionally, because they are bathed by CSF, they can also sense the presence of toxins in CSF.
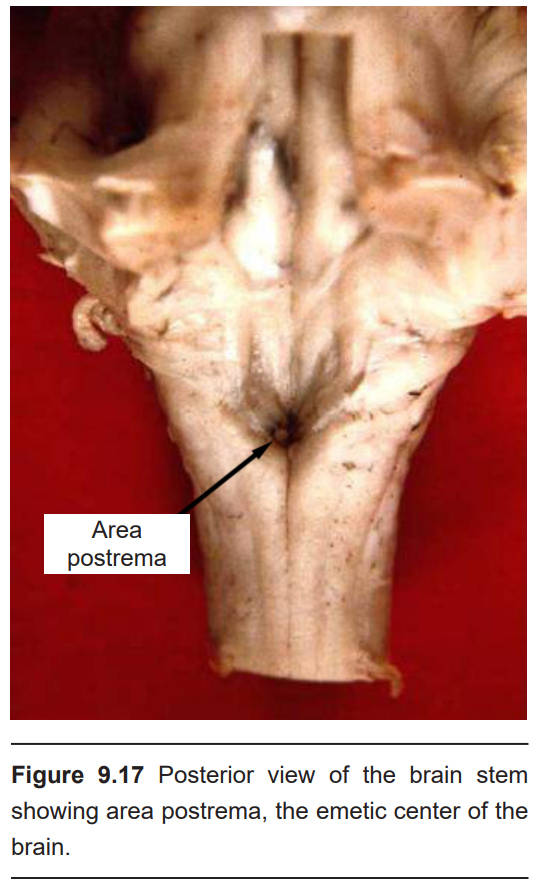
Image Credits
Cover: https://pixabay.com/photos/pork-ribs-dinner-pork-food-meat-2157179/
9.1 https://commons.wikimedia.org/wiki/File:Southern_spotted_skunk.jpg
9.2 https://commons.wikimedia.org/wiki/File:1543,Vesalius%27OlfactoryBulbs.jpg https://commons.wikimedia.org/wiki/File:Anatomy_of_the_woodchuck_(Marmota_monax)_(2005)_(18007398570).jpg olfactory structures outlined by Austin Lim
9.3 https://commons.wikimedia.org/wiki/File:Head_olfactory_nerve.jpg
9.4 https://commons.wikimedia.org/wiki/File:Location_of_olfactory_ensheathing_cells_(OECs)_within_the_olfactory_system.png modified by Austin Lim
9.5 https://commons.wikimedia.org/wiki/File:Cribriform_plate_Close-up_view.png
9.6 https://commons.wikimedia.org/wiki/File:Olfactory_Sensory_Neurons_innervating_Olfactory_Glomeruli.jpg modified by Austin Lim
9.7 https://pixabay.com/photos/grain-bread-bread-rye-bread-cut-3135224/
9.8 https://commons.wikimedia.org/wiki/File:Entorhinal_-_DK_ATLAS.png
9.9 https://commons.wikimedia.org/wiki/File:MRI_of_orbitofrontal_cortex.jpg
9.10 https://commons.wikimedia.org/wiki/File:Anatomy_and_physiology_of_animals_Taste_buds_on_the_tongue.jpg modified by Austin Lim
9.11 https://commons.wikimedia.org/wiki/File:1402_The_Tongue.jpg modified by Austin Lim
9.12 https://openbooks.lib.msu.edu/neuroscience/chapter/taste-central-processing/ modified by Austin Lim
9.14 ttps://commons.wikimedia.org/wiki/File:EMA401_Mechanism_of_Action.jpg
9.15 https://commons.wikimedia.org/wiki/File:Gray806.png phrenic nerve highlighted by Austin Lim
9.15 ttps://commons.wikimedia.org/wiki/File:Buffer_Part_1.png modified by Austin Lim
9.17 https://commons.wikimedia.org/wiki/File:Human_caudal_brainstem_posterior_view_description.JPG modified by Austin Lim
The Open Neuroscience Initiative is funded by a grant from the Vincentian Endowment Fund of DePaul University.
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License.
