8 Chapter 8: Sensation and Perception – Other Physical Senses
Cover Image
Austin Lim, PhD (DePaul University)
Editor: Alexander Rajan, PhD
Our nervous system is equipped with a variety of specialized biological “tools” that can detect much more than just photons of light. We can detect the shape of air waves, and interpreting those signals gives us sound information and the perception of music. We can detect stimuli with our skin, such as temperature, pressure, and textures. We can also detect physical information about our own bodies, as in the way our head is tilted or the position of our limbs. In this chapter, we describe these other forms of physical stimuli, and how that information is conveyed to and represented in the brain.
All sensory systems, including vision (Chapter 7) and the systems described below, follow the same general path of communication into our nervous systems and awareness. First, the incoming signal must reach a cell (generally called a transducer) that can change its electrical properties in response to the stimulus (such as a retinal photoreceptor cell responding to photons). Then, that information initiates a series of signals into the CNS, reaching structures such as the thalamus (in most cases; olfaction being an exception), primary sensory cortical areas, and finally, higher order perception. Although there are several sensory components throughout our body that detect these signals, there are no sensory receptors in our central nervous system.
As we discuss sensory and perception further, it’s good to keep in mind that not all sensations are perceived. For example, your body can detect changes in blood pressure, and your brain responds to these changes activating compensatory neural circuits. However, this response does not reach consciousness and is not perceived.
Chapter 8 outline
8.1 The Auditory System
8.2 The Vestibular System
8.3 The Somatosensory System
8.1 The Auditory System
Unlike photons of light, sound waves are compressions and rarefactions of a medium. For us land animals, that medium is usually air, but sound waves can propagate very well in water or through solids. Without a medium, sound fails to propagate: As the tagline from the thriller Alien reads, “In space, no one can hear you scream.”
8.1.1 Sound waves
Before we get to the anatomical structures involved in sound perception, it is important to first understand the physical nature of sound waves. All sounds, from the clattering of a dropped metal pan to the melodies of a Mozart violin concerto, are contained in their corresponding sound waves. There are three components of sound waves.
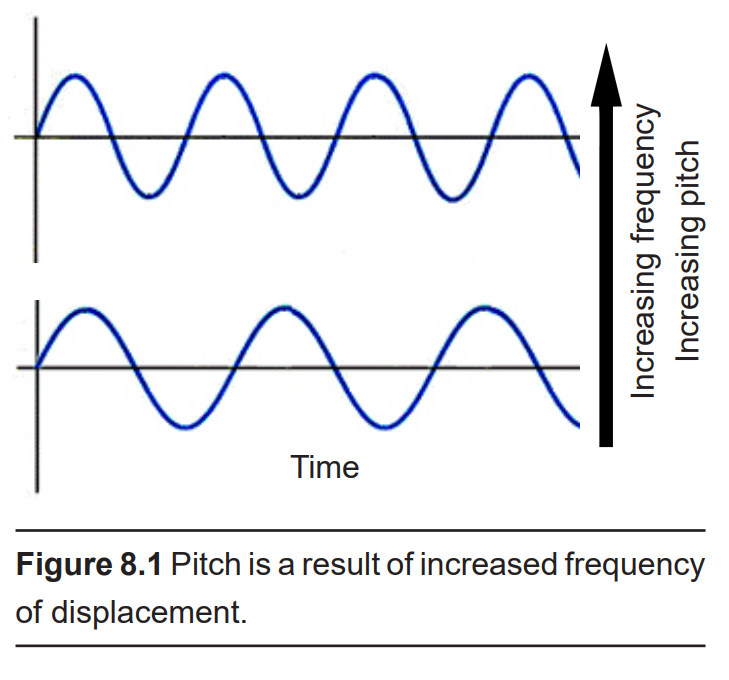
Frequency, or “How often do the sound waves compress?”
The more often they repeat, the higher the pitch. The highest notes humans are able to hear is around 20,000 Hz, a painfully-shrill sound for those who can hear it. People often tend to lose their high frequency hearing as they age. On the opposite end of the spectrum, low frequency sounds are the deep rumbles of bass, and the human ear can hear sounds down in the 20 Hz range. Doubling the frequency changes the tone by an octave: concert A is 440 Hz, and the A an octave higher is 880 Hz.
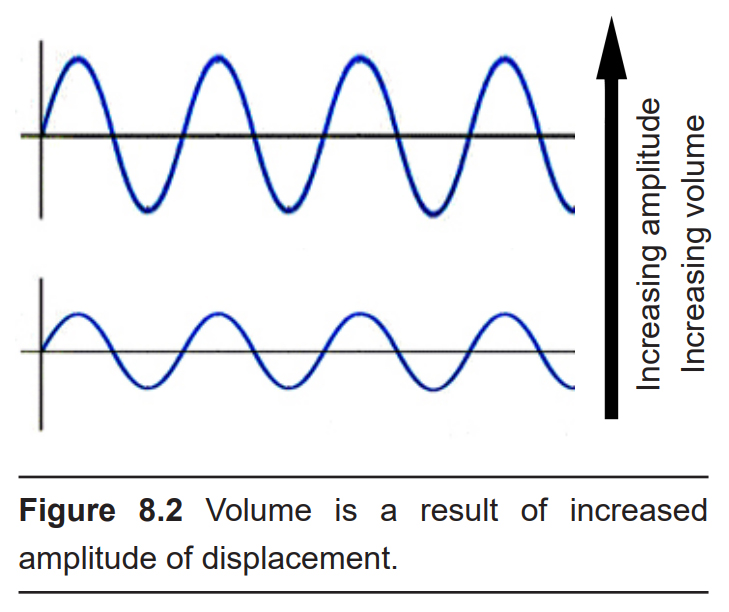
Amplitude, or “How much do the waves displace the medium from baseline?”
The larger the amplitude of the wave, or the greater distance between the peak and the trough of the signal, the louder the sound is. Loudness is measured in decibels (dB). To give you an idea of approximate sound intensities, the background noise of a quiet library is about 40 dB, and a typical conversation is close to 60 dB. A rock concert or lawnmower is between 100 and 110 dB, which is right around the pain threshold. Prolonged exposure to these high amplitude sound waves can lead to permanent damage to the auditory system resulting in hearing loss or tinnitus (a ringing in the ear, even in the absence of a sound stimulus).
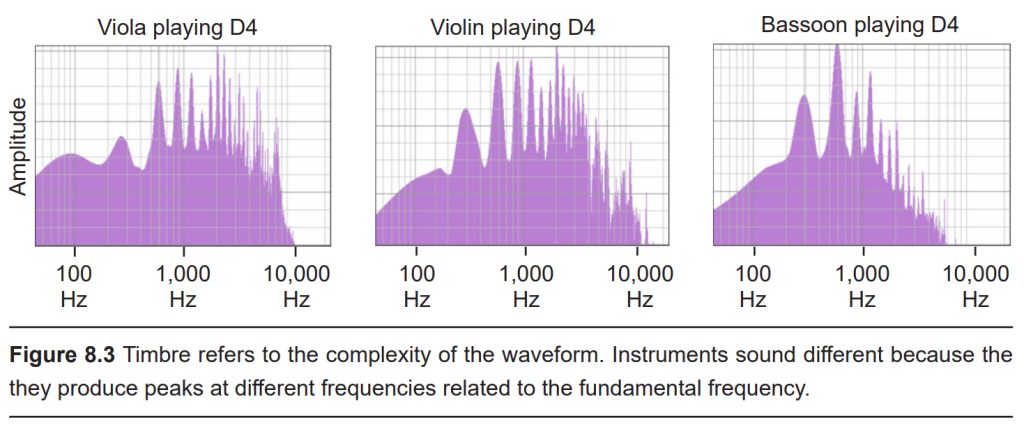
Timbre, or “How complex is the wave form?”
Timbre (TAM-bur) refers to the “color” or the “character” of the sound. Both a piano and a guitar could play the exact same note, yet these two instruments would produce very different sounds. This happens because the sound waves created by each instrument differ in complexity. Both notes contain the same fundamental frequency which gives us the pitch of that note. But each wave differs in the number of other high frequency components called overtones that are contained within that note. These overtones oscillate at a frequency that are multiples of the fundamental frequency (twice as often, for example). A sound wave without any overtone frequencies is called a pure tone, or a sine wave tone.
8.1.2 Physical structures of the auditory system
Our auditory system is a series of physical structures and nervous system components that are responsible for conveying sound waves into meaning and context.
The external component of the auditory system begins with the pinna or auricle, or more simply, the ear. Its shape functions as a funnel, capturing and channeling sound
waves into the auditory canal. The pinna and the auditory canal are parts of the outer ear. Also, because the pinna is asymmetrical, its shape helps us determine where a sound is coming from. In some nonhumans, the pinna serves these functions and more: some animals are able to disperse excess heat through their ears (elephants), and some even use them to display emotion (dogs, horses).
At the end of the auditory canal is the tympanic membrane, or ear drum. This membrane is a very delicate piece of tissue at only 0.1 mm thin, and is subject to damage by physical injury such as head trauma, nearby explosions, or even changes in air pressure during scuba diving. When incoming sound waves reach the tympanic membrane, it vibrates at a matching frequency, amplitude, and timbre. The tympanic membrane also represents the boundary between the outer ear and the middle ear.
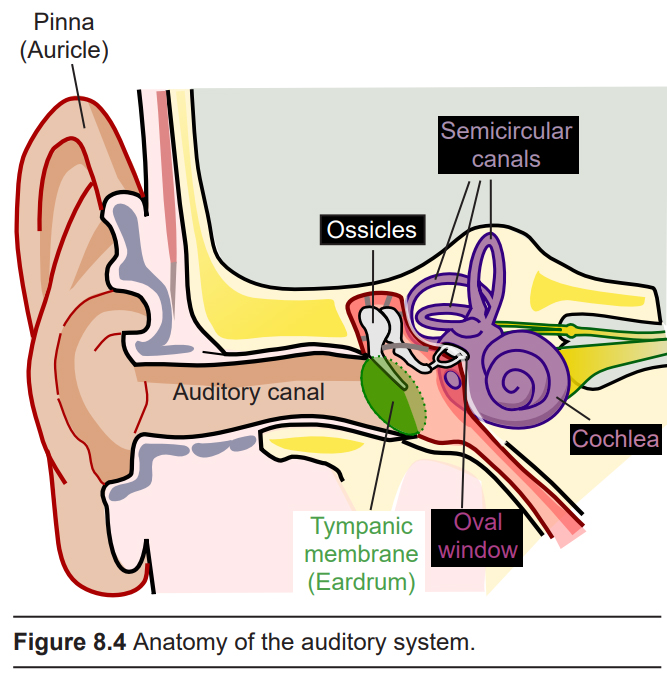
Physically attached to the tympanic membrane are the ossicles, a series of three bones that convey that vibrational sound information. These bones in order, called the malleus, incus, and stapes, conduct vibrations of the tympanic membrane through the air-filled middle ear. The tympanic membrane and the ossicles function to amplify incoming sounds, generally by a tenfold difference. This amplification is important because the inner ear is filled with liquid rather than air, and sound waves do not travel very well when moving from air into a denser medium – think about how muffled sounds are when you submerge your head underwater.
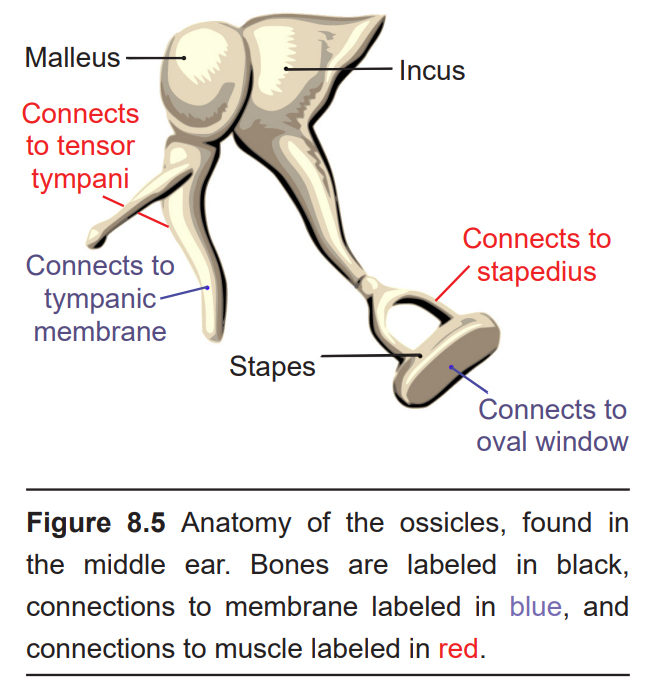
The movement of the ossicles is partially regulated by two different muscles, the tensor tympani muscle which connects with the malleus, and the stapedius muscle which connects to the stapes. When these muscles contract, the ossicles move less, which decreases the intensity of loud sounds. This response, called the acoustic reflex, dampens incoming sound by about 15 dB. (This is why we talk much louder than normal when we first leave a concert: we have lessened auditory feedback from our ears, so we tend to talk louder to compensate.) Crickets use a similar reflex so that they don’t deafen themselves while chirping!
The stapes, the third of the ossicles, is physically connected to the oval window, a thin membrane that is at the entrance to the main auditory structure of the inner ear, a hollow bone called the vestibular labyrinth. The auditory part of this structure is the spiral-shaped cochlea (cochlea is named for the Ancient Greek word “snail shell”.) Think of the cochlea as a rolled-up cone that makes about 2 and 3/4 turns. If this cone was theoretically unrolled, the widest diameter portion (called the base) would be closest to the oval window, while the narrowest portion (called the apex) would be at the center of the spiral.
This change in shape from the base to the apex is important: objects with different stiffness vibrate at different frequencies. The base of the cochlea is stiffer than the apex, and so it vibrates are higher frequencies than the apex. This makes different points along the spiral of the cochlea responsive to different frequencies.
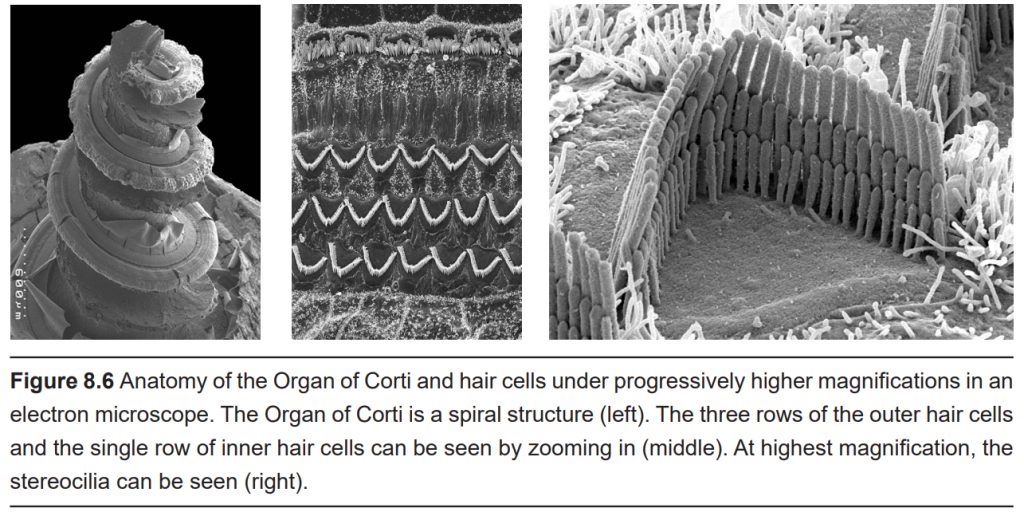
Inside the cochlea is a specialized epithelial membrane called the Organ of Corti. The Organ of Corti is the first nervous system structure that is responsible for processing physical vibrations and converting them into signals that the nervous system can interpret.
8.1.3 Neural components of the auditory system
The Organ of Corti contains the components necessary for converting sound waves into action potentials. Adjacent to the Organ of Corti in the cochlea is a liquid called endolymph. The endolymph is a high potassium, low sodium solution that is similar to CSF. It makes up the extracellular solution in the inner ear.
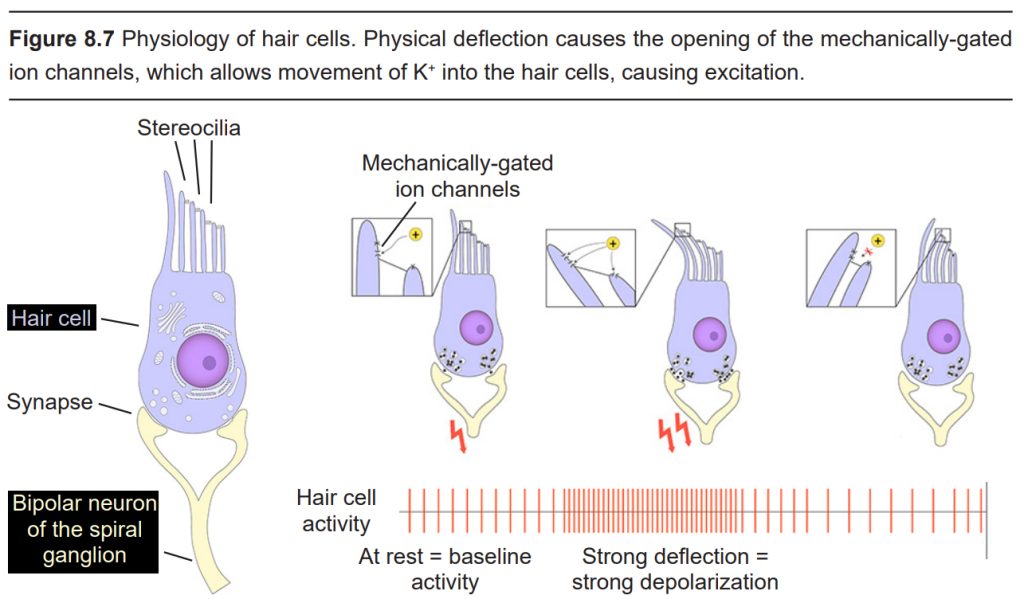
Embedded along the interior surface of the Organ of Corti are the somata of hair cells, the primary sensory neurons that interpret physical movement. They are named “hair cells” because of their cellular structure; each hair cell has somewhere between 30 and a few hundred hair- shaped stereocilia that protrude away from the Organ of Corti, reaching into the endolymph. We have two different populations of hair cells, the inner hair cells and outer hair cells.
Movement of the inner hair cells is important for the detection of sounds. When a vibration reaches the oval window, the endolymph also vibrates, causing a physical movement of the membranes within the cochlea, which displaces the stereocilia. The physical movement causes mechanically-gated ion channels to open, allowing K+ to enter the hair cells, resulting in depolarization and neurotransmitter release. This process of physical motion that leads to neural signaling is called mechanotransduction. Hair cells are very sensitive to stereocilia movement: a deflection on the order of 100s of picometers (a
picometer is one trillionths of a meter) is sufficient to cause changes in electrical signaling in a hair cell!
The length of the stereocilia differs between hair cells, and these differences allow them to respond to different frequencies of vibrations. Shorter hair cells are more responsive to high pitch sounds, and are located closer to the base of the cochlea. On the other hand, longer hair cells are more sensitive to lower pitched sounds, and are located near the apex, in the center of the spiral. Accordingly, there are two “tuning” mechanisms in the cochlea that make specific inner hair cells sensitive to specific frequencies: the position along the spiral, and the length of the stereocilia.
The outer hair cells function as an amplifier to increase the intensity of vibrations. It is estimated that the outer hair cells increase sound by anywhere between 20 and 80 dB. We have three rows of outer hair cells.
Clinical connection: Hearing loss
Many people experience permanent hearing loss, a decrease in volume by 25 dB or more. Hearing loss is divided into two categories. Conductive hearing loss is a result of changes to the auditory system up to the oval window, such as a tumor in the ear canal, a perforation of the tympanic membrane, or changes in middle ear pressure (such as how everything sounds muffled while changing altitudes when an airplane takes off, for example). Sensorineural hearing loss results from changes at the level of the inner ear or further up in the neural pathway, such as hair cell damage, a brain tumor, bacterial or viral infections, or exposure to various toxins or drugs.
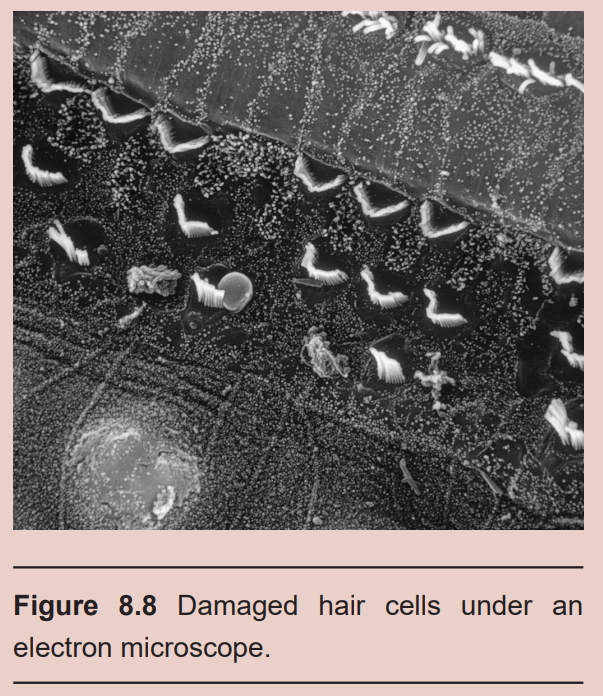
The most common cause of hearing loss is excessive noise exposure. Although the acoustic reflex is capable of dampening the intensity of the incoming vibrations, prolonged exposure to high amplitude sound waves can still cause damage. Motorcycles, the maximum volume on headphones, or loud venues like concerts and clubs can produce sounds in the 95-110 dB range, which can cause some permanent hearing loss. Additionally, the acoustic reflex is not fast enough to minimize damage from sudden, loud sounds in excess of 120 dB, such as a gunshot. All these sources of acoustic trauma are preventable by wearing appropriate hearing protection, which can decrease the intensity of sounds by up to 30 dB.
Old age is another common cause of hearing loss, likely because older people have had more accumulated exposure to noise. An estimated 1 in 3 people older than 65 have hearing deficits. We are born with about 15,000 hair cells, but throughout the course of our life, many get damaged irreparably. The shorter hair cells are more sensitive to injury, so it is common for people to lose sensitivity to high-frequency sounds. The loss of these hair cells can begin as early as a person’s 20s – some places intentionally play high frequency sounds to prevent teenagers from loitering.
Partial hearing loss can be reversed with the help of medical devices. A hearing aid is a processor that helps to filter out background noise, decrease pitch, and amplify incoming sounds. A cochlear implant is a surgically- implanted device that receives incoming sound information and directly stimulates the auditory nerve via electrodes, bypassing the external components of the auditory system.
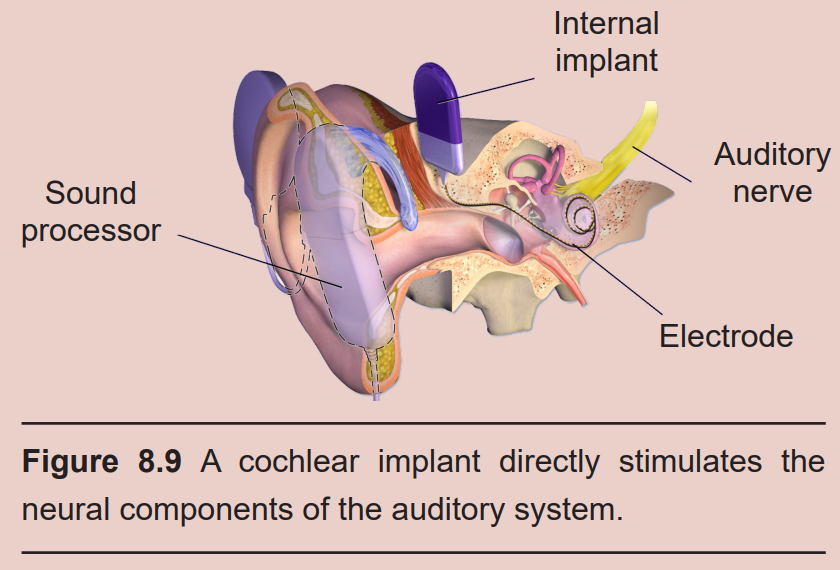
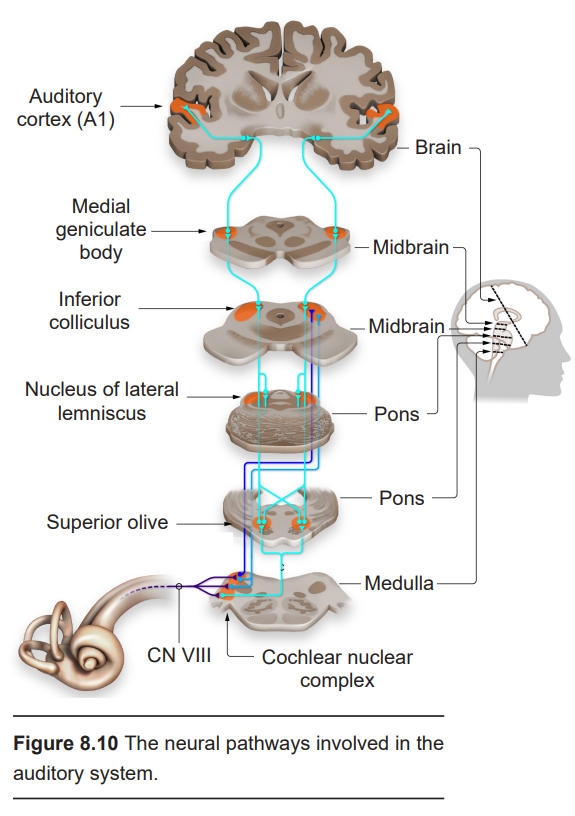
Hair cells form glutamatergic synapses onto bipolar neurons of the spiral ganglion. The axons of these neurons make up the vestibulocochlear nerve (Cranial nerve VIII, historically called the auditory nerve. This nerve also carries vestibular information, which is also processed in the inner ear. See section 8.2.2 for more details.) Some neurons project into a pontine area called the superior olive, an integrative center that receives bilateral inputs from both ears. Other neurons project into an area of the rostral medulla called the cochlear nuclear complex, which carries out some auditory processing functions.
One of the primary functions of the superior olive is to help us figure out if a sound originates from the left or the right side of our head. To do this, the superior olive performs two different calculations. First, sounds are louder the closer you are to them. A difference in volume between what one ear and the other ear perceives, called the interaural level difference, is evaluated by neurons in the lateral superior olive. The second calculation is related to the speed of sound. Sound travels roughly a kilometer in three seconds. Because of this delay, sounds reach one ear sooner than it reaches the other. This interaural time difference is assessed by neurons of the medial superior olive. The ability for these neurons to properly localize sound is the result of years of training.
Because both ears (in humans and most non-humans) are at the same height, sounds above and below can be difficult to localize. This is why sometimes people and animals tilt their heads to try to hear better. Some species of owls have ear openings at different heights on their heads, increasing their ability to localize the source of a sound on the vertical axis.
Auditory information is then passed into inferior colliculus (IC). Signaling within the IC is important for interactions between multiple sensory inputs and a motor response. These IC neurons are particularly responsive to biologically-relevant sounds, such as unexpected noises, which may signal an approaching predator. Processing in the IC helps the animal focus their attention towards these stimuli.
The IC then conveys that auditory information into the medial geniculate body, one of the nuclei of the thalamus. These thalamic neurons then send projections into the primary auditory cortex, or A1 (historically called Herschel’s gyrus.) A1 is the dorsal- most part of the temporal lobe.
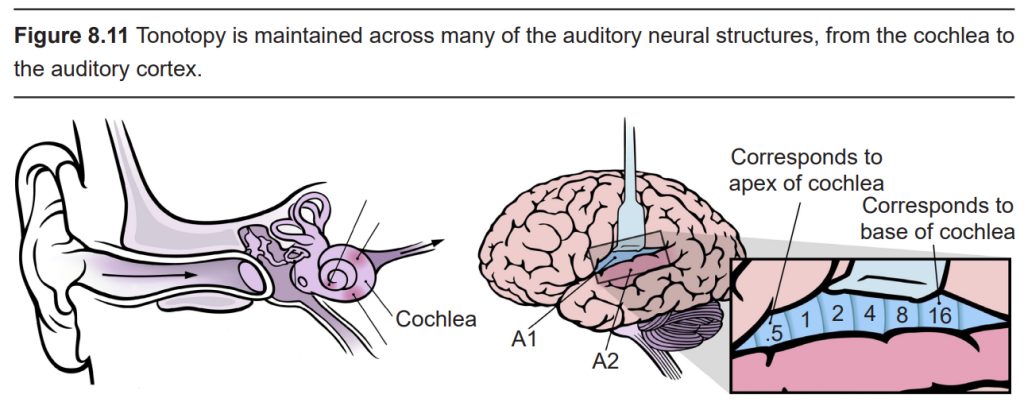
Many of the neural processing components of sound, such as the Organ of Corti, the spiral ganglion, and A1, are tonotopically organized, meaning that adjacent physical areas are responsible for conveying information from adjacent frequencies. For example, the hair cells that respond most to 440 Hz vibrations are right next to cells that respond maximally to 441 Hz, but far away from cells that respond most to 14,000 Hz. Likewise, in the cortex at A1, the cells that best process 440 Hz are adjacent to those that best process 441 Hz, but far away from those that maximally respond to 14,000 Hz.
After signal processing in A1, that sound information is then passed through the secondary and tertiary auditory cortices, which likely carry out higher order auditory functions. One of the earliest studied cortical areas involved in auditory processing is Wernicke’s area, which is critical to the understanding of speech and language (See chapter 14 for more details).
There is evidence to support a dual stream organization to auditory perception, just like in the visual system in cortex. The dorsal auditory stream helps identify the location of sounds (analogous to the where pathway), speech production, and language related memory, while the ventral auditory stream contributes to sound recognition (similar to the what pathway) and sentence comprehension.
Clinical connection: Tinnitus
Tinnitus is a condition characterized by the occasional perception of a ringing, whistling, buzzing, or clicking sound in the absence of a genuine stimulus. Tinnitus is a symptom rather than a disease. It often appears after some degree of hearing loss or injury; alternatively, it can come and go without any particular underlying cause. An estimated 15% of people experience some amount of tinnitus, but about 2% of people have clinically significant tinnitus.
It is still unknown what causes tinnitus, however a common predictor of tinnitus is hearing loss. This correlation has led researchers to our current leading theory about tinnitus, which suggests that the perception of phantom sounds originates in the brain as a result of faulty plasticity: the “understimulated” brain, which doesn’t receive expected auditory inputs, produces these abnormal signals.
8.2 The Vestibular System
When we tilt our head to the side, or look up and down, that movement information is conveyed to our brain using the vestibular system. The vestibular system is a sort of three- dimensional compass that can detect head movement, and that information helps us figure out how our head is oriented and how to balance ourselves in changing conditions.
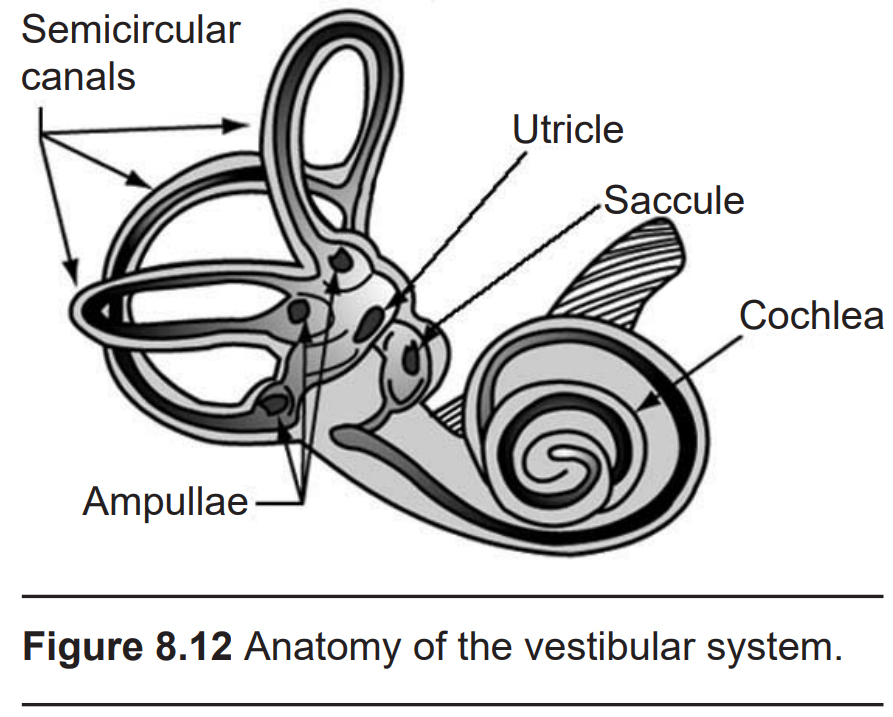
The vestibular system is made up of two structures that are intimately tied in with the anatomical features of the inner ear.
8.2.1 The Otolith Organs
Next to the cochlea and within the vestibular labyrinth are two membranous sacs, the saccule and the utricle. Collectively, these structures are responsible for determining changes in inertia. The saccule is more sensitive to vertical movements, like when you are standing in a moving elevator. The utricle is more responsive to horizontal movements, such as when driving around a corner too quickly.
Within the saccule and utricle are otoliths, a series of small calcium carbonate crystals (the prefix oto– meaning “ear”, and the suffix –lith meaning “rock”) embedded in a 50 uM thick gelatinous membrane. Also embedded within this membrane are the stereocilia of a different population of hair cells. These otolith hair cells are biologically similar to the hair cells found in the cochlea: deflection of the stereocilia allows for K+ in the surrounding endolymph to enter the cells through mechanically-gated ion channels. The main difference is the nature of the stimulus that cause the sterocilia to bend. In the cochlea, vibrations in the surrounding tissue cause hair cell movement. In the saccule and utricle, however, it is a shifting of the physical weight of the otoconia that result in hair cell movement.
The information encoded by the hair cells is passed into the brain via a branch of the vestibulocochlear nerve (CN VIII). These axons send projections to several brain areas, notably the cerebellum, which is a structure critically important for balance. CN VIII also projects into the reticular formation of the brain stem, the spinal cord, and the thalamus.
8.2.2 The Semicircular Canals
The semicircular canals are the structures that are responsible for detecting head rotation. Anatomically, they are a series of three arch-shaped membranous tubes within the vestibular labyrinth, each one oriented at a right angle to each other. Because of this shape, the semicircular canals sense and convey information about any direction of head movement: roll, pitch, and yaw.
These semicircular canals are filled with endolymph, the same potassium-rich solution that is in the cochlea that is important for auditory sensation. At the end of each of the three canals is a small swelling called the ampulla. Contained in the ampulla is a gelatinous membrane called the cupula. Here, hair cells extend stereocilia, as well as one cellular protrusion called a kinocilium, into the cupula. When we tilt our head, the endolymph in the semicircular canals flows in the ampulla, which physically displaces cilia. As in the auditory system, these hair cells have mechanically-gated ion channels which work on a “push-pull” system: when the stereocilia are deflected in one direction, the hair cells depolarize, while deflection in the opposite direction causes hyperpolarization.
Clinical connection: Vertigo
Vertigo is the sensation of spinning or movement while standing still. Vertigo often leads to dizziness, imbalance, ear pain, nausea, or vomiting. It can indirectly lead to injuries, especially as a person stands up or if they experience vertigo while driving. A symptom rather than a disease itself, an estimated 7% of people experience vertigo in their lifetimes, affecting women about 3 times as often as men.
There are a variety of conditions that could cause a person to experience vertigo. Benign paroxysmal positional vertigo can come and go spontaneously and is generally not a sign of an underlying health condition. People with Meniere’s disease, a chronic condition diagnosed in early adulthood, often experience vertigo along with other ear-related symptoms such as tinnitus or hearing loss. Bacterial or viral infections can cause inflammation of the inner ear, resulting in abnormal vestibular signaling. Excessive alcohol intoxication decreases the density of the endolymph and potentiates inhibitory signaling, resulting in exaggerated motor responses following head movement. Severe head trauma that damages the inner ear may also cause vertigo.
Vestibular reflexes
Many axons from the pontine vestibular areas send projections into the cerebellum, and these signals are important for reflexes related to balance. For example, imagine you are standing facing forward on a crowded bus when it stops abruptly. The sudden change in inertia causes your body to reflexively pushing your toes into the ground, preventing your body from toppling forward. This behavior is driven by neural signaling in the vestibular organs and their communication with the cerebellum.
A similar postural reflex that depends on vestibular inputs is the righting reflex. This is a behavior that develops early in several animals from humans to Drosophila, and it allows the animal to correct their body position if they are in an abnormal orientation (imagine a baby who can get back to crawling on all fours after they have fallen on their side). Generally, this reflex is learned during the first few weeks of life. Knowing up from down depends on the afferent vestibular signals, however performance of the motor task also requires integration of visual and somatosensory inputs.
A more subtle reflex is the vestibulo-ocular reflex (VOR), which is observed as our eyes stay fixated on a target while our head moves. This allows our vision to stay focused, even as the head is moving. For example, after rotating your head to the right, the eyes reflexively move to the left, which allows for the visual field to be stabilized briefly. It can be observed as a physician performs a diagnostic assessment called the rapid head impulse test, where they move your head quickly side to side (as if you were shaking your head “no”) while watching your gaze.
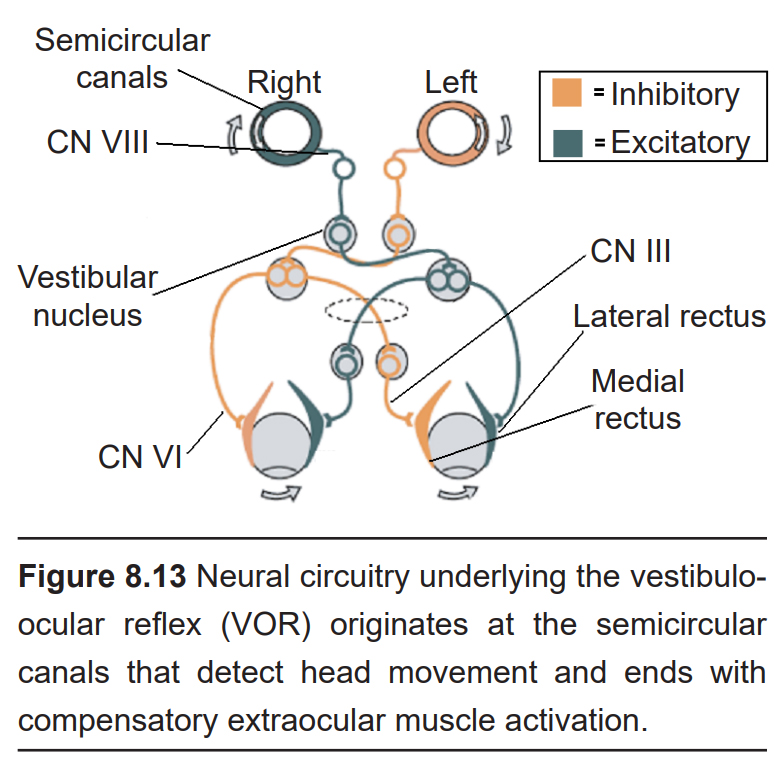
This response is driven through a series of three synapses. The first synapse is formed between the axonal projections from neurons of the vestibular system, and neurons of the vestibular brainstem nuclei. From here, these neurons send axonal projections to the contralateral hemisphere (that is to say, their axons decussate) and form synaptic connections with two populations of neurons in the contralateral pons. One set of motor neurons excite the extraocular muscle opposite of the eye movement: a right head turn will trigger excitation of the lateral rectus of the left eyeball, which pulls the left eyeball in the temporal direction (left). The other population are interneurons that eventually excite the medial rectus of the right eyeball, the extraocular muscle that pulls the right eyeball in the nasal direction (left, again). Simultaneously, there are inhibitory circuits that act at the opposite muscles to inhibit the eye from moving in the same direction as the head turn. Operating on the span of about 10 ms, the VOR is one of the fastest reflexes in the body.
8.3 The Somatosensory System
The nerves that receive somatosensory inputs (soma– referring to the body, in the same way the cell body is also called the soma) are the afferent branch of the somatic nervous system, a component of the peripheral nervous system. They give us the ability to detect information about the body. These nerves can sense a wide variety of physical stimuli, including pressure, stretch, vibration, heat, and pain.
The information about our body gets processed minimally in spinal cord circuits, ascends through the brain stem, passes through signal processing within the thalamus before reaching circuits in the somatosensory cortices. The first cortical region to receive these signals is the primary somatosensory cortex, or S1, which is the anterior-most gyrus of the parietal lobe, immediately adjacent to the central sulcus. In primates, S1 is subdivided into Brodmann areas 3a, 3b, 1, and 2.
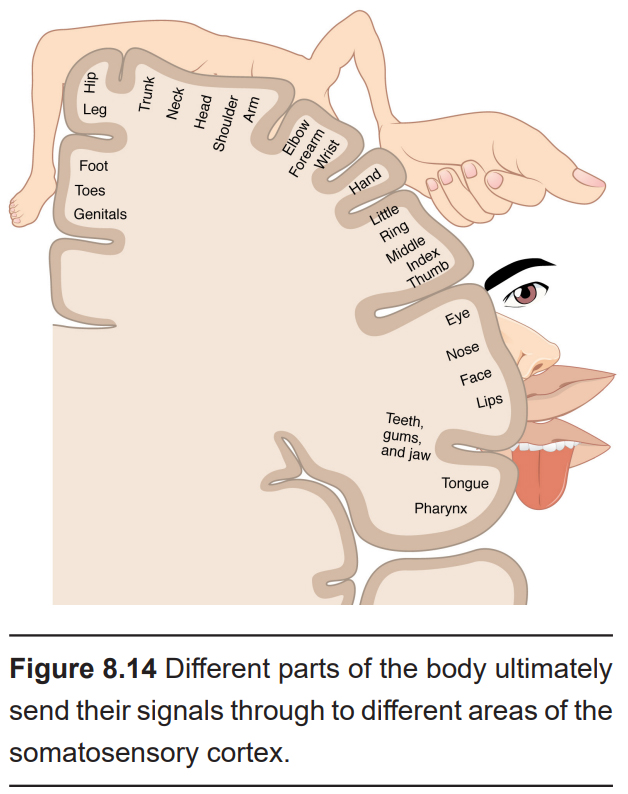
There are many different types of sensory receptors in the skin, each of which detects a very specific type of stimulus: for example, deep movement of joints, vibrations as you run a finger over a fabric, or the cold touch of an icicle in winter. For some perceptions, many of these sensory receptors are combined, but often these sources of information are carried in separate but parallel paths, a theory called the labelled line principle. The type of information (e.g., light touch on your finger) can be tracked from the sensory receptor, through specific parts of the spinal cord, through nuclei of the thalamus, and into particular subregions of primary somatosensory cortex.
The neurons in the spinal cord, thalamus, and S1 demonstrate somatotopic organization, meaning that specific points on the body map to specific neural populations. For example, touching the skin on the hip will activate neurons found in the dorsal-most aspect of S1, while sensations on the tongue activate the neurons of ventral and lateral S1. Thus, there are two key bits of information kept together throughout the somatosensory pathway: what kind of stimulus, and where.
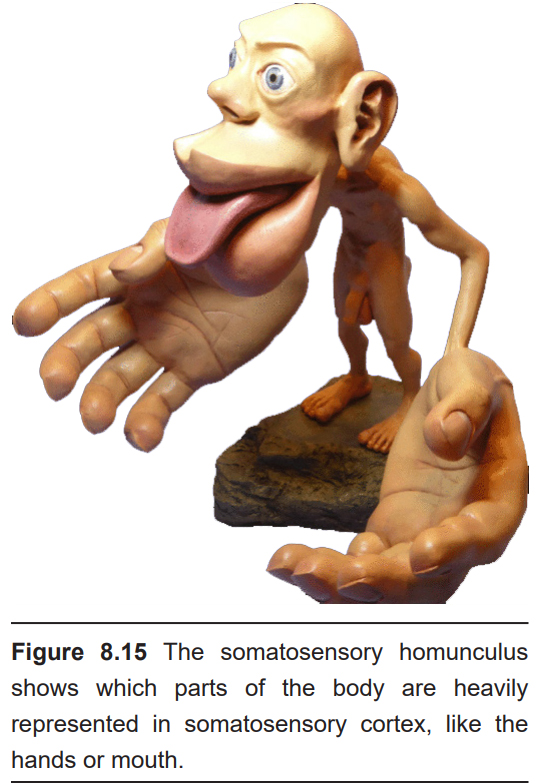
The mapping of this organization was discovered during brain surgery. In epilepsy surgery, parts of the operation are performed as the person is awake. The famed neurosurgeon Wilder Penfield electrically stimulated a small section of S1, then asked the person what they felt. Patients would respond that they feel sensations in their hand when the medial S1 was activated, the little finger when a more ventral area was activated, and so on. In repeating this process while moving slowly across the span of S1, Penfield mapped out the organization of S1. One key observation in doing this process was the volume of brain regions corresponding to different body parts was not necessarily proportional to the volume of skin: for example, a large portion of the cortex was dedicated to sensory information from the lips or the fingertips, while very little was dedicated to the shoulders or trunk. This observation indicates that certain areas of the body are more densely packed (lips, fingertips) with sensory neurons compared to others (shoulders, trunk). A representation of a person drawn to the scale of how large each area is represented in S1 is called the sensory homunculus.
Some of the outputs of S1 then project into the secondary somatosensory cortex, or S2, located posteriorly within the parietal lobe, where higher order processing takes place.
8.3.1 Cutaneous receptors
Imagine reaching your hand into your pocket. In doing so, you may feel something circular with tiny ridges all along the perimeter, roughly an inch in diameter. A series of highly specialized sensory receptors just under the surface of our skin (the “cutis” in cutaneous refers to the skin) allows us to sense this stimulus. That information travels through ascending, afferent nerves into S1 and S2, and this is where we perceive that we have a quarter in our pocket.
It helps to divide the variety of cutaneous receptors into three categories, roughly based on the types of sensations they detect and convey.
Mechanoreceptors
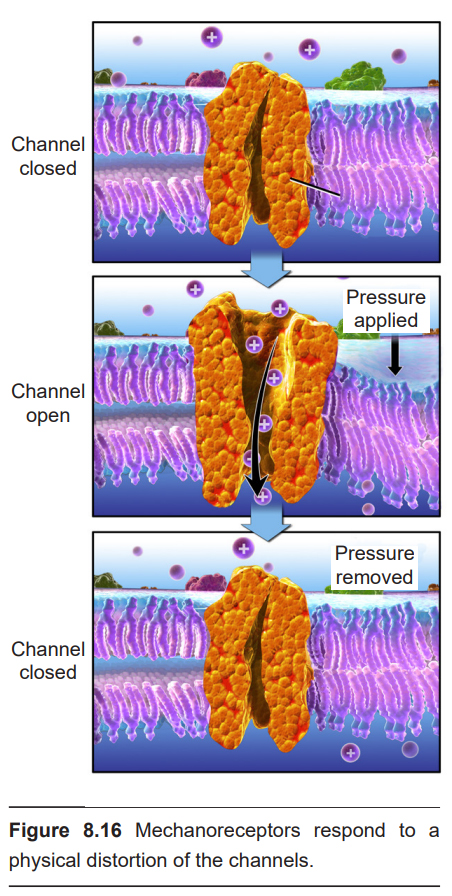
Cutaneous (meaning found in the skin) mechanoreceptors are responsible for sensing mechanical changes to the skin, such as pressure or stretch. On a molecular scale, these mechanoreceptors detect changes at the skin using mechanically-gated ion channels. These transmembrane proteins are specialized for detecting the physical distortion of the channel, similar to the channels found in the stereocilia of the hair cells or vestibular cells. When pressure is applied to these proteins, the cation channel opens and Na+ moves down its electrochemical gradient into the neuron, causing depolarization.
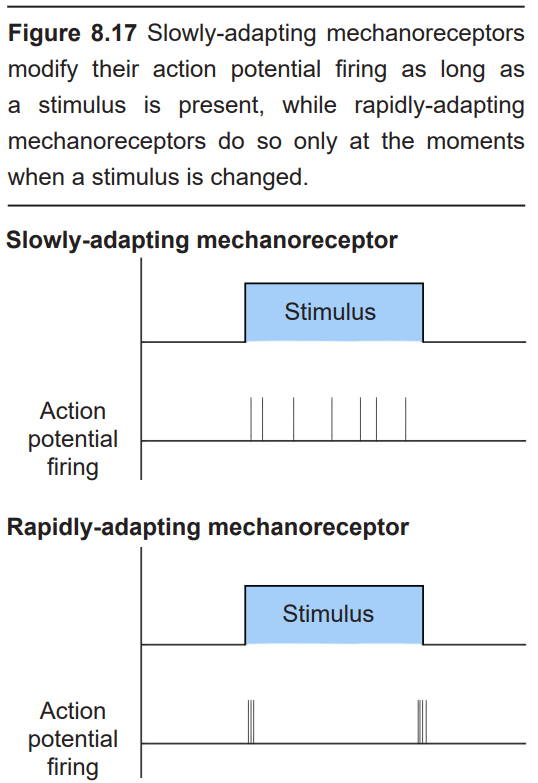
The class of mechanoreceptors can be divided into two categories based on their electrical properties in response to stimuli: slowly- adapting and rapidly-adapting. Slowly-adapting mechanoreceptors change their action potential firing rate as long as the stimulus is present, while rapidly-adapting mechanoreceptors only change activity at the moment there is a change in stimulus. For example, imagine the stimulus of a coin that is sitting in the palm of your hand. The slowly-adapting sensory neurons may increase their firing rate as long as the coin is in your palm. When the quarter is removed, they return to their baseline firing rate. These are sometimes also called tonic receptors. However, a rapidly- adapting sensory neuron will change its action potential firing properties only at two timepoints: the moment the coin lands in your hand, and the moment that coin is removed. These neurons signal a change in status and are sometimes also called phasic receptors.
There are four different classes of cutaneous mechanoreceptors, two of which are slowly-adapting and two which are rapidly- adapting. These different mechanoreceptors have slightly different shapes, and are therefore capable of detecting different types of stimuli.
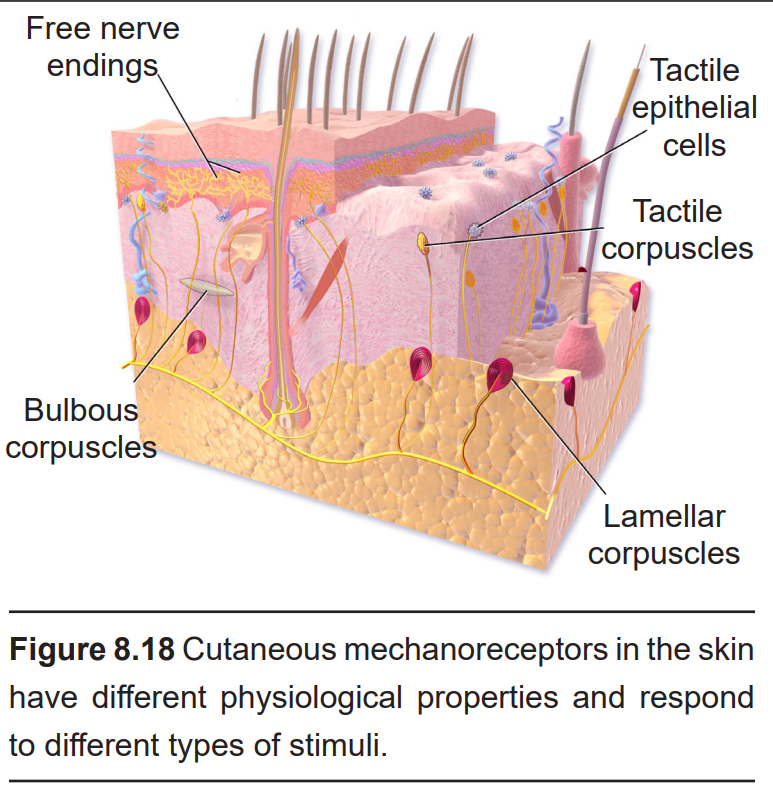
- Tactile epithelial cells (also called Merkel’s discs) are located in the superficial skin layers, and are very densely populated underneath our fingertips. They sense pressure and help us perceive edges, points, and corners. These are the most precise mechanoreceptors with resolution at the level of 0.5 mm, which allow us to detect different distances in bumps while running your fingers across Braille. These tactile epithelial cells are slowly-adapting receptors, and they release serotonin at their synapses.
- Lamellar corpuscles (also called Pacinian corpuscles) are mechanoreceptors wrapped in several layers of connective tissue. They mostly respond to high-frequency vibrations and deep pressure. They are rapidly-adapting receptors and are the deepest of the cutaneous receptors.
- Tactile corpuscles (also called Meissner’s corpuscles) are highly sensitive to light touch, skin movement, and low-frequency vibration. Like the tactile epithelial cells, tactile corpuscles are also found in the superficial skin layers, concentrated heavily at the fingertips. These receptors are rapidly-adapting receptors.
- Bulbous corpuscles (or Ruffini endings) respond to stretching of the skin, such as the sensation of an object slipping out of a closed hand, for example. These bulbous corpuscles are slowly-adapting receptors.
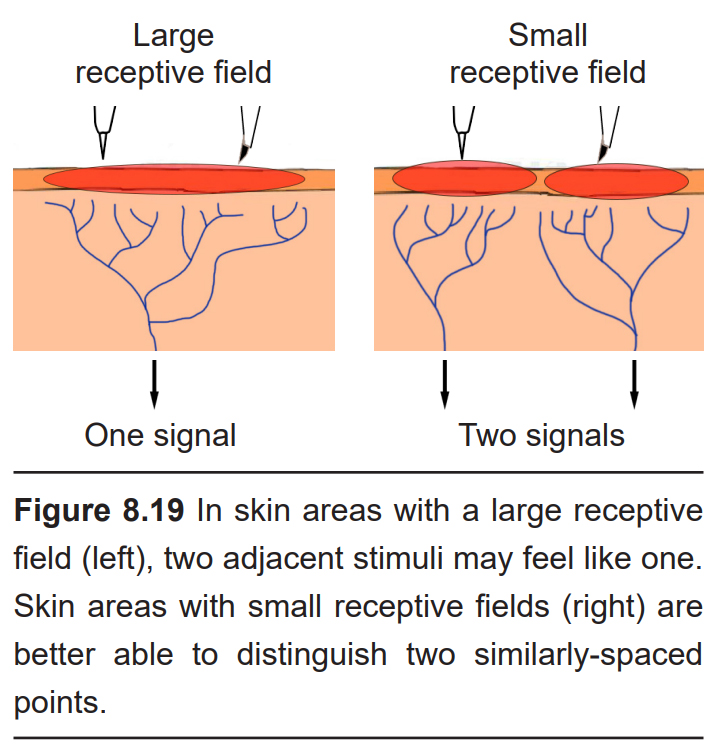
Each cutaneous mechanoreceptor is able to detect somatosensory information in an area of skin. The particular patch of skin that responds to particular cutaneous stimuli is a receptive field, and the smaller the receptive field, the better the brain can distinguish between two different adjacent tactile stimuli. Receptive fields differ in size throughout the body. For example, the receptive fields at the fingertips and the lips are very small (roughly 10 mm2), while the receptive fields on the back are much larger. Additionally, different classes of mechanoreceptors have different sizes of receptive fields: deeper structures, like lamellar corpuscles, have larger receptive fields compared to receptors found closer to the skin surface.
The approximate size of a receptive field can be assessed in people using a two-point discrimination task. Here, the experimenter will place two adjacent stimuli simultaneously onto the skin of a blindfolded person at different places. The patient is then asked if they feel one or two stimuli. In areas with large receptive fields, like the top of the thigh for instance, a pair of points 2.5 cm apart (1 inch) may feel like one single point of contact. The distance between the two points is increased incrementally until the patient reports they first feel 2 points.
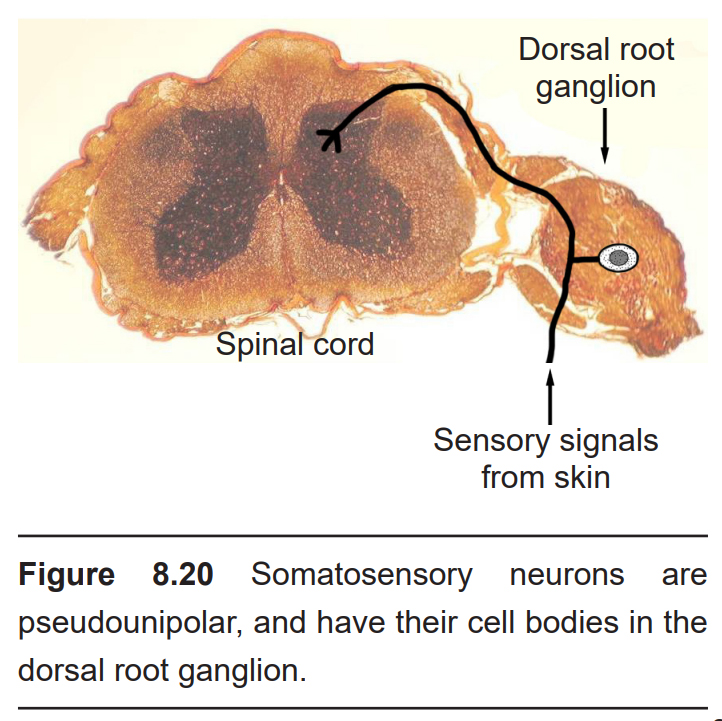
The somatosensory afferents from these receptors have their cell bodies located in the dorsal root ganglion (DRG), a clump of somata that are close to the dorsal side of the spinal cord. These touch receptors are the primary neurons, and they are classified as pseudounipolar neurons based on their morphology. Pseudounipolar neurons don’t have true dendrites; instead, each soma sends a single protrusion which branches into two directions, one towards the skin surface where it terminates as one of the mechanoreceptors, and the other direction towards the spinal cord. Mechanosensory information travels via Aβ axons, which are large diameter (~10 μM) axons that can send signals on the order of 50 meters per second.
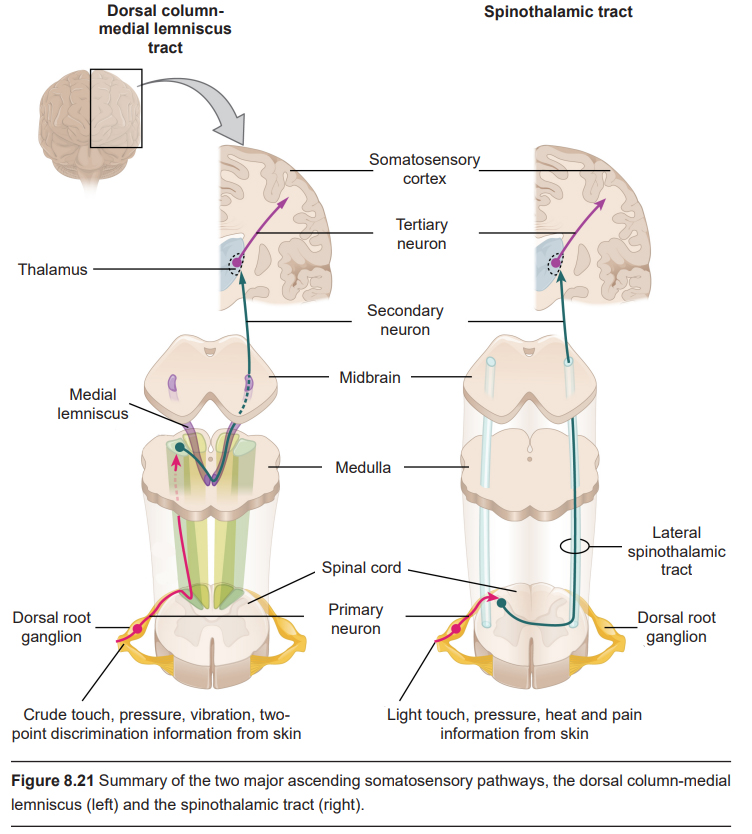
The next neurons in the ascending signaling pathway are the secondary neurons of the spinal cord, which have their cell bodies in the dorsal horn. These neurons send ascending axonal projections through the two different signaling pathways. The dorsal-column medial lemniscus (DCML) tract sends projections through the ipsilateral white matter. These DCML signals carry crude touch, pressure, vibration, and two-point discrimination information. The other ascending pathway is the spinothalamic tract, which projects upwards through the contralateral white matter. Spinothalamic tract carries light touch and pressure information, as well as heat and pain (described below).
The DCML neurons form synapses onto neurons found in the medulla. At this point, these neurons decussate (cross the midline), then ascend further into the ventral posterior lateral (VPL) nucleus of the thalamus. Spinothalamic tract neurons project directly into the thalamus.
The thalamic neurons are the tertiary neurons of the somatosensory system. They project into S1, in particular Brodmann areas 3b and 1.
Thermoreceptors
Thermoreceptors are the cutaneous receptors that sense temperature. There are two classes of thermoreceptors. The low-threshold thermoreceptors detect innocuous, non-harmful temperatures in the range of 15 to 45 degrees C. High-threshold thermoreceptors detect painful and potentially damaging temperatures hotter than 45 C or colder than 15 C.
Thermoreceptors are located in a class of cutaneous receptors called free nerve endings. Compared to the mechanoreceptors, free nerve endings are found closest to the surface of the skin. Like mechanoreceptors, the free nerve endings are the sensory end of the pseudounipolar neurons which have their cell somata in the dorsal root ganglion.
Expressed on these free nerve endings are a population of receptors called transient receptor potential receptors (TRPchannels). All proteins are sensitive to changes in temperature, but these TRP channels change shape more dramatically than other proteins, making them ideal for sensing temperature. TRP channels are non-selective cation channels. Cold sensations are detected by the action of the TRPM8 receptor, which is also activated by menthol, a chemical isolated from peppermint that produces a cooling sensation. Warm sensations are sensed by the TRPV1 receptor, which is also activated by the chemicals capsaicin, the compound that makes spicy peppers feel hot, and allyl isothiocyanate, which is found in wasabi.
Temperature information does not get passed along the same neurons that carry mechanosensitivity signals. Cool and cold temperature sensation is passed through thinly- myelinated Aδ fibers, which are smaller in diameter (~5 μM) and transmit signals slower (~25 m/s) compared to the Abeta fibers. Warm and hot temperature sensation are passed through unmyelinated C fibers, which are even smaller (~1 μm) with slower conduction velocity (~1 m/s).
The secondary neurons of the thermoreceptors are within the dorsal horn of the spinal cord, and the ascending pathway runs through the white matter of the lateral aspect of the spinothalamic tract.
Nociceptors
Detection and avoidance of pain is a highly-adaptive behavior that can improve the odds of survival chances of an animal. Pain detection is carried out by nociceptors. Nociceptors can detect a variety of noxious stimuli, ranging from crush to acid and high heat. They are expressed on free nerve endings. Many pain-detecting neurons respond to more than one type of noxious stimulus, and these are called polymodal nociceptors. Nociceptors also send their signals through Aδ and C fibers. Adelta fibers detect sharp, highly-localized pain, while C fibers carry a more dull, throbbing pain that is difficult to pinpoint.
Clinical connection: Pain disorders
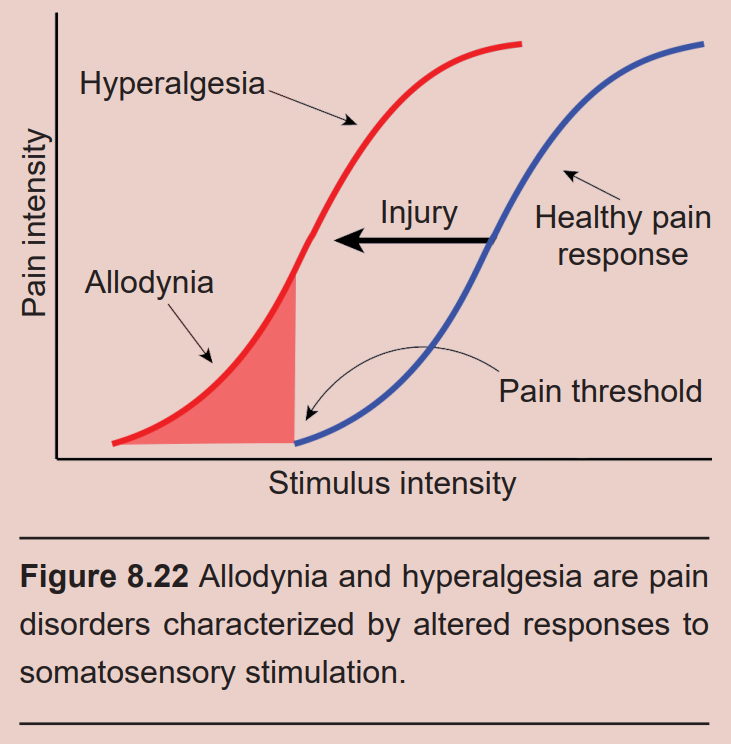
In healthy individuals, nociceptors that detect pain send those signals into the spinal cord. But in people experiencing allodynia, non-injurious tactile stimuli, such as strands of hair brushing across your forehead or the feeling of a shirt resting on your shoulders, cause the sensation of pain. Diabetes, physical trauma, or postherpetic neuralgia may cause allodynia. Drugs that modify the action potential firing properties of the afferent somatosensory pathway, such as topical anesthetics (voltage- gated sodium channel inhibitors) or calcium channel antagonists, can help relieve the pain.
A related disorder is hyperalgesia, an abnormally heightened perception of pain. Hyperalgesia can be the result of sensitization or as a withdrawal symptom of opioid use. Both allodynia and hyperalgesia can be symptoms of neuropathic pain, a broad category of pain conditions resulting from damage to the nervous system.
People experiencing pain even in the absence of pain-producing stimuli may be diagnosed with chronic pain. Chronic pain may be localized or widespread, affecting large areas of the body at once. Up to 10% of the global population may experience chronic pain, and a variety of risk factors contribute, from advanced age, being male, low socioeconomic status, unhealthy lifestyles, and a history of surgical intervention. Chronic pain can lead to significantly shorter lifespans and may contribute to the opioid misuse epidemic.
A philosophically-opposite condition is called congenital insensitivity to pain (CIP). People with CIP are incapable of perceiving pain, no matter how severe. CIP is a rare genetic condition that is observed from birth. People with CIP have significantly shorter lifespans since they do not sense that their bodies may be injured, and may not learn to avoid exposure to damaging stimuli. Some patients may walk for days on a compound fracture of the shin bone, or never even notice that their back is broken. CIP is associated with mutations in the SCN9A gene, which codes for a component of the voltage-gated sodium channels that are expressed in nociceptors.
There are several molecular components that make up the pain detection systems in the skin.
Some mechanoreceptors are likely expressed on free nerve endings. Nociceptive mechanoreceptors can sense crushing, shearing, or cutting of the skin. These may have a high threshold of activity, which prevents them from being activated under harmless circumstances.
Acid-sensing ion channels are cation channels that respond to low pH conditions in the dermis, which is usually seen in inflammation, which is often downstream of tissue injury. The inflammation causes a release of various cellular signaling molecules (such as prostaglandins and cytokines) which cause sensitization, an enhancement of future incoming pain stimuli.
High-threshold thermoreceptors are also involved in pain detection. On a molecular level, the same TRP channels that sense cool and hot (TRPM8 and TRPV1) are also implicated in detecting painfully cold or painfully hot temperatures.
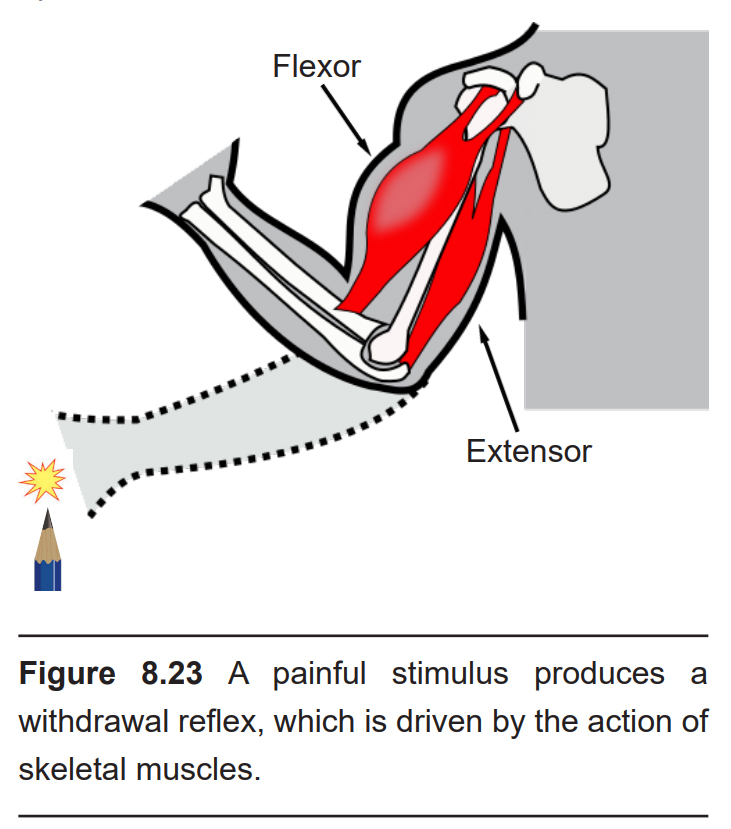
Imagine you are reaching into your bookbag and accidentally stick your fingertip with the sharp end of a pencil. Within half a second, the withdrawal reflex causes a series of muscular changes which moves your hand away from the pointy end. This motor response is driven by a circuit of neurons in the spinal cord, and it is mediated completely independently from descending motor control from the brain. The nociceptive input from the sensory neurons enters the spinal cord through the dorsal horn where it forms a synapse onto an excitatory interneuron. This interneuron then signals to two other populations. The first is the motor neuron that innervates the flexor, a generic term used to describe muscles that withdraw your hand when contracted. The other connection is formed with an inhibitory interneuron, which innervates the motor neuron that controls the extensor, a generic term used to describe the muscle that functions opposite the flexor. In total, simultaneous activation of an extensor and inhibition of the extensor result in the rapid withdrawal of the hand.
In addition to communicating with the neurons involved in the withdrawal reflex, the nociceptive signals also send ascending projections through the white matter of the lateral spinothalamic tract.
Clinical Connection: Referred pain
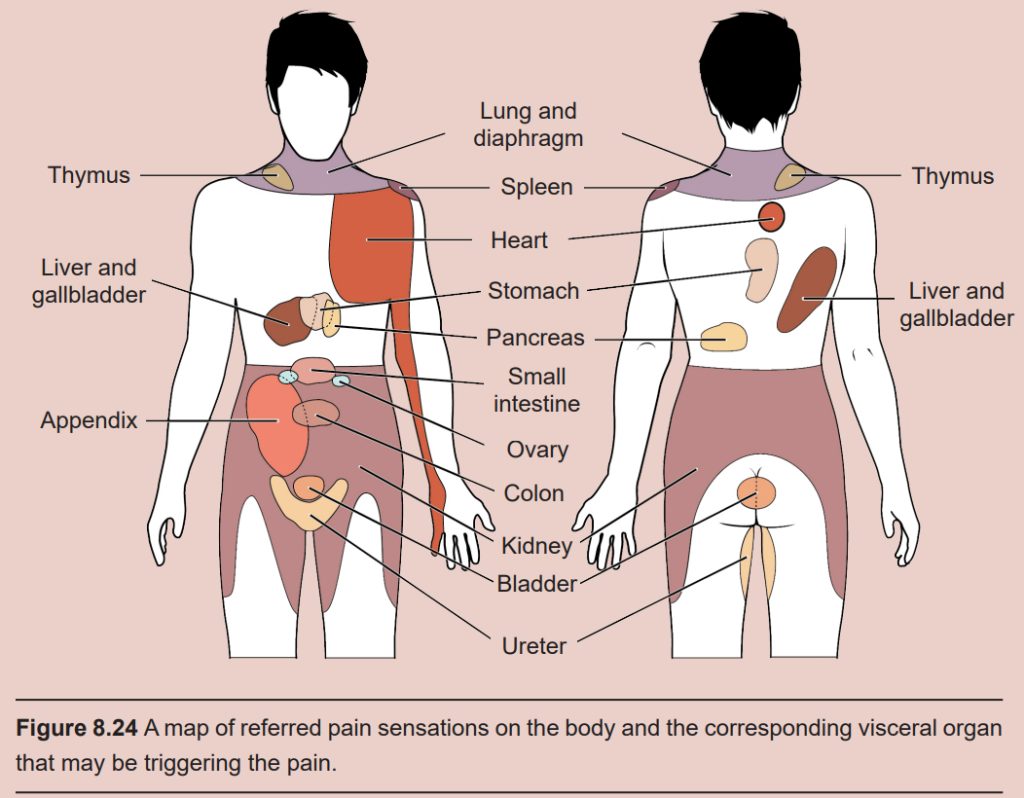
One of the warning signs that a person is having a heart attack is perception of pain in the shoulders or medial aspect of the left arm. This sensation is referred pain: the feeling of pain at a site separate from where an injury is located. Referred pain happens when the nervous system is unclear about how to process signals from an internal organ, and the brain interprets those afferent signals as bodily pain. Injury at different internal organs cause different patterns of somatic pain: liver injury may present as pain in the shoulder blades, lung cancer sometimes causes shoulder pain, and kidney stones can cause pain in the lower back, abdomen, and sides. Even the headache-like pain you get from a “brain freeze” from drinking cold liquids too quickly is a form of referred pain.
It is unclear what causes referred pain. The convergence projection theory suggests that the afferent projections from the internal organs and the nociceptive somatosensory neurons of the skin form synapses onto the same population of spinothalamic tract secondary neurons. When these secondary neurons project towards the brain, there is no ability to differentiate the origin of the signal coming from the primary neuron. The central sensitization theory suggests that the neurons of the spinal cord change in their excitability threshold with prolonged exposure to injurious stimuli. So, when an injured internal organ repeatedly sends signals into the secondary neurons, a host of neurotransmitters cause sensitization of the nociceptive signaling, which are now activated under non-noxious conditions.
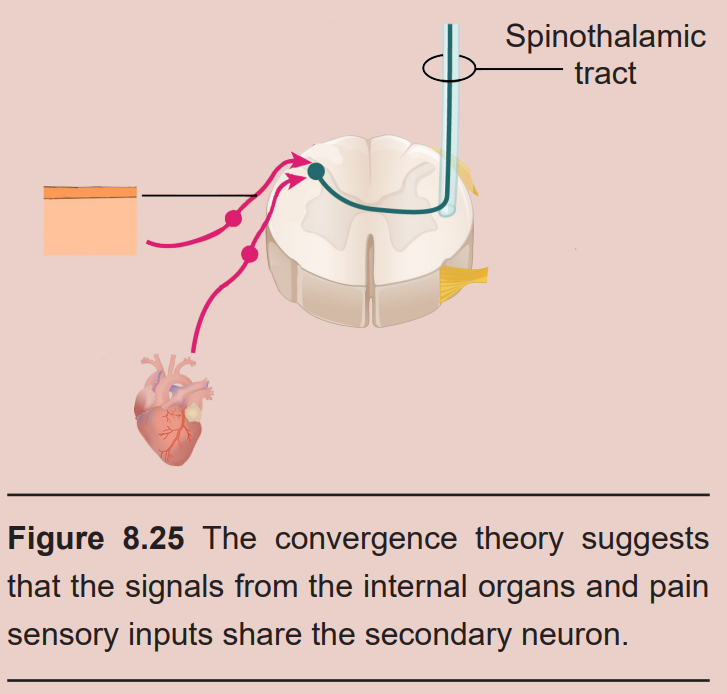
8.3.2 Proprioception
Raise your arms above your head. Even without seeing your arms, your nervous system has mechanisms that inform you about the location and position of your body parts, including how much your joints are bent. This sense is called proprioception and is critically important for coordinated movement and motor reflexes that contribute to those tiny, rapid adjustments that are made while maintaining balance.
Proprioceptive information ascends through the spinal cord and into the brain via the dorsal column-medial lemniscus tract. Proprioception is also processed in S1, more specifically Brodmann areas 3a and 2.
There are two main neural systems that work together to give us our sense of proprioception.
Muscle spindles
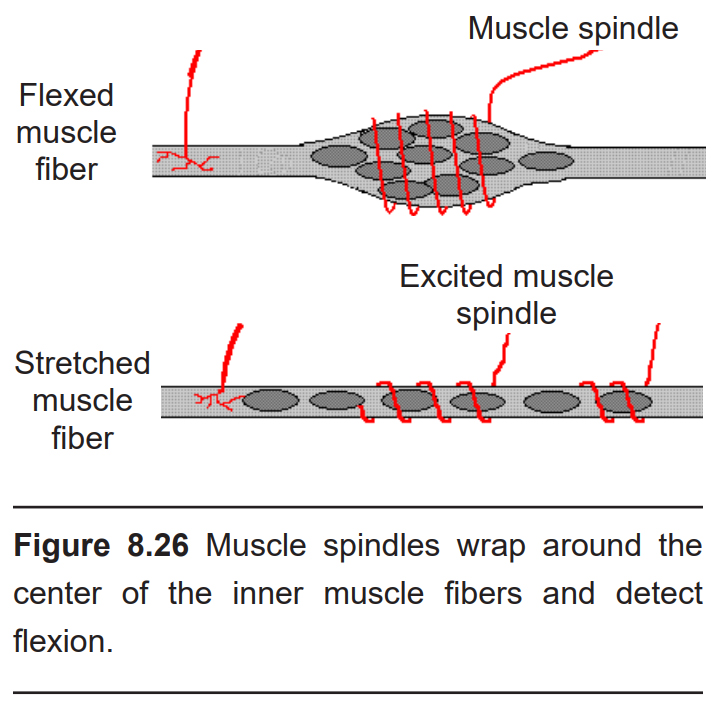
Wrapped around the intrafusal skeletal muscle fibers is a series of nervous structures called muscle spindles which detect the status of the muscle. Each muscle spindle is between 6 and 10 mm long, and spirals around the thickest part of the muscle fiber. When a skeletal muscle is flexed, it widens in diameter, becoming shorter and thicker. As it does, the muscle spindle wrapped around it also changes in shape. The muscle spindles communicate this information to the nervous system through the action of nonselective-cation mechanoreceptors that respond to physical distortion – muscle stretching increases firing rate, while muscle flexion decreases firing rate. Muscle spindle density is much greater among skeletal muscles that are used for very precise movements (like in the hand) compared to those used for coarse movements (like in the thighs).
There is also a motor function of these muscle spindles. Through a circuit of neurons in the spinal cord, they communicate with the gamma motoneurons that terminate at these intrafusal muscle fibers. The combination of the sensory and motor components allows the muscle spindles to function in the stretch reflex (also called the myotatic reflex), a spinal cord- mediated response to muscle stretch that causes flexion to prevent excess stretching, which can cause damage to the muscles.
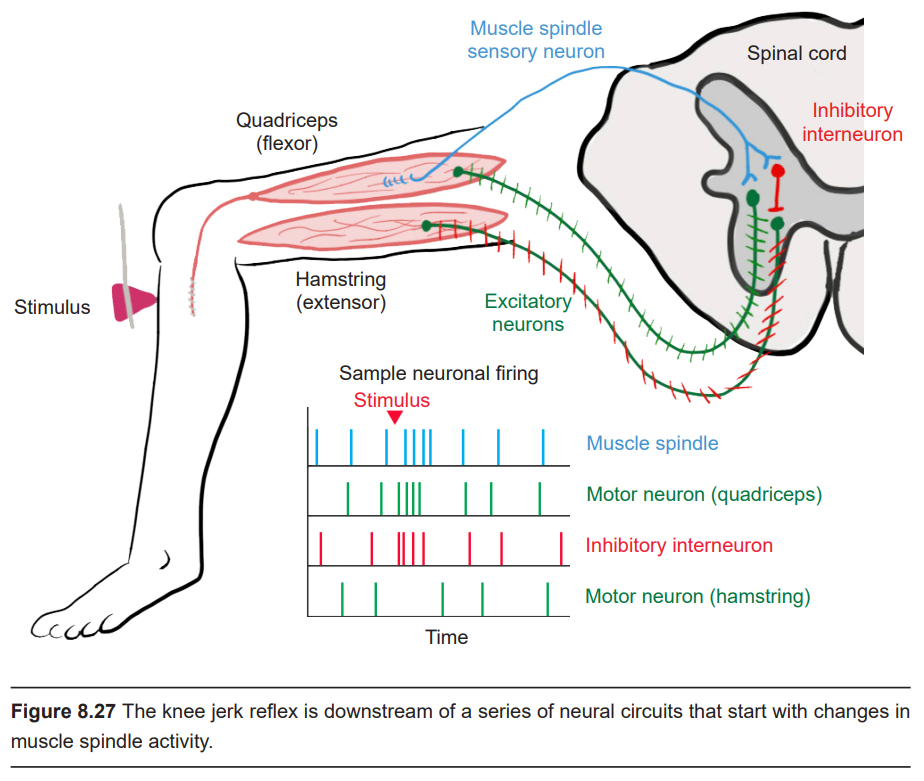
The most-well known example of the stretch reflex is the knee-jerk reflex (or the patellar tendon reflex), which may be tested in a standard medical physical. Here, the patient sits on the end of the exam table with their knee bent and their lower leg dangling freely over the edge. The examiner uses a pointed rubber hammer to gently tap on the tendon that connects the kneecap (patella) to the muscle in front of the shin bone. When tapped, it causes a stretch of the quadriceps, the muscle on the top of the thigh. The muscle spindles detect this extension of the muscle, they communicate this information with an excitatory motor neuron of the spinal cord that causes a flexion of the quadriceps, which causes the foot to kick forward. The circuitry of this reflex is described as a monosynaptic reflex arc. (There is also a spinal cord inhibitory interneuron that receives muscle spindle inputs that decreases the activity of the hamstring, the muscle on the bottom of the thigh. In this reflex, the quadriceps is the flexor while the hamstring is the extensor.)
The neural circuitry responsible for the knee-jerk reflex is located in the L2, L3, and L4 regions of the spinal cord. If the knee-jerk reflex is weak or absent, there may be some degeneration or death of the nerves that communicate with the muscles, or possibly an injury to these specific areas of the spinal cord.
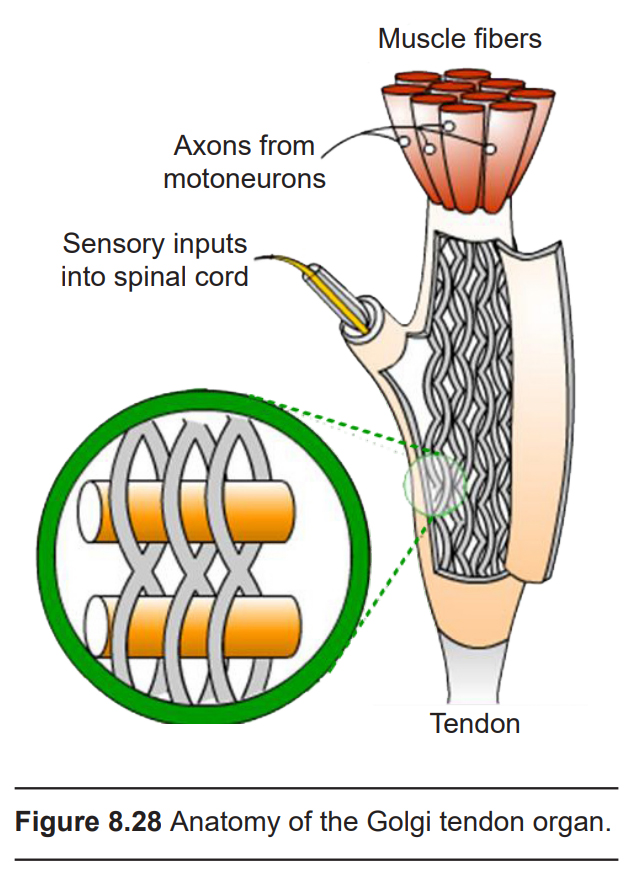
Golgi tendon organs (GTOs)
Made up of collagen fibers, GTOs are found at the insertion site between muscles and tendons. Signals from the GTO convey information about the amount of the tension that each set of skeletal muscles is experiencing as we move around. They also contribute to our detection of weight, as we lift something heavy for example. Each GTO connects to about 20 muscle fibers and is about 0.5 mm long.
During muscle contraction, the tension in the GTO increases. Contained within the GTO are nonselective cation mechanoreceptors that open during this physical deformation (similar to those found in other aspects of the somatosensory system) causing changes in the excitable properties of the GTO. The outputs of the GTO communicate with interneurons found in the spinal cord. These interneurons in turn inhibit the motor neurons that innervate the muscle that is “pulling” on the tendons.
The GTO communicates via Aα fibers. These large diameter axons (15 μM) are heavily myelinated, and can transmit action potentials as fast as 100 meters per second, which are the fastest projections in the somatosensory system.
The Open Neuroscience Initiative is funded by a grant from the Vincentian Endowment Fund of DePaul University.
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License.
Image credits
Cover: https://pixabay.com/photos/guitar-ropes-instrument-music-wood-56916/
8.1 https://upload.wikimedia.org/wikipedia/commons/1/18/Sound_wave_physics.gif Modified by Austin Lim
8.3 https://commons.wikimedia.org/wiki/File:Frame-level_Instrument_Recognition_by_Timbre_and_Pitch.pdf
8.4 https://upload.wikimedia.org/wikipedia/commons/e/ee/Anatomy_of_the_Human_Ear_blank.svg
8.5 https://commons.wikimedia.org/wiki/File:Auditory_ossicles-en.svg
8.6 https://wellcomecollection.org/works/fsg372tb https://wellcomecollection.org/works/jfsbzy4g
8.7 https://upload.wikimedia.org/wikipedia/commons/2/2c/Hair_cell_action.png
8.8 https://wellcomecollection.org/works/hrvtw6fx
8.9 https://commons.wikimedia.org/wiki/File:Cochlear_Implant.png modified by Austin Lim
8.10 https://commons.wikimedia.org/wiki/File:Auditory_Pathway.png modified by Austin Lim
8.11 https://commons.wikimedia.org/wiki/File:Frequency_mapping_in_human_ear_and_brain_-_10.1371_journal.pbio.0030137.g001-L.jpg modified by Austin Lim
8.12 https://commons.wikimedia.org/wiki/File:Vestibular_organs-_canals,_otolith,_cochlea.jpg modified by Austin Lim
8.13 https://commons.wikimedia.org/wiki/File:Vestibulo-ocular_reflex.PNG modified by Austin Lim
8.14 ttps://commons.wikimedia.org/wiki/File:1421_Sensory_Homunculus.jpg
8.15 ttps://commons.wikimedia.org/wiki/File:Front_of_Sensory_Homunculus.gif
8.16 https://commons.wikimedia.org/wiki/File:Mechanically_Gated_Channel.png modified by Austin Lim
8.18 https://commons.wikimedia.org/wiki/File:Blausen_0809_Skin_TactileReceptors.png modified by Austin Lim
8.19 https://commons.wikimedia.org/wiki/File:Compass_tool.svg modified by Austin Lim https://commons.wikimedia.org/wiki/File:Skin_numbers.jpg modified by Austin Lim
8.20 https://commons.wikimedia.org/wiki/File:Histological_preparation_of_DRG.jpg; https://commons.wikimedia.org/wiki/File:Structure_of_sensory_system_(4_models)_E.PNG modified by Austin Lim
8.21 https://commons.wikimedia.org/wiki/File:1417_Ascending_Pathways_of_Spinal_Cord.jpg modified by Austin Lim
8.22 https://commons.wikimedia.org/wiki/File:Hyperalgesia_and_allodynia.tif modified by Austin Lim
8.23 https://commons.wikimedia.org/w/index.php?curid=53328677 https://pixabay.com/vectors/pencil-education-office-stationery-23648/ modified by Austin Lim
8.24 https://commons.wikimedia.org/wiki/File:1506_Referred_Pain_Chart.jpg
8.25 https://commons.wikimedia.org/wiki/File:1417_Ascending_Pathways_of_Spinal_Cord.jpg modified by Austin Lim; https://commons.wikimedia.org/wiki/File:Human_Heart.png
8.26 https://commons.wikimedia.org/wiki/File:Spindle.GIF
8.27 ttps://commons.wikimedia.org/wiki/File:Knee_jerk_reflex.png
8.28 https://commons.wikimedia.org/wiki/File:Tendon_organ_model.jpg
