7 Chapter 7: Sensation and Perception – The Visual System
Cover Image
Austin Lim, PhD (DePaul University)
Editors: Dana Simmons, PhD (University of Chicago)
Allison Morales, OD (Illinois College of Optometry)
At over 400 miles of explored passages, the Mammoth Cave system in Kentucky is the longest known cave system on Earth. During the cave tour, the guides momentarily extinguish all the lights. The void that follows is overwhelmingly eerie. Opening and closing your eyes makes no difference. You may have difficulty figuring out which way is up and down, and you might even lose your balance when you tilt your head.
Humans are remarkably dependent on the visual system to gain information about our surroundings. Consider how tentatively you walk from the light switch to your bed right after turning off the lights!
The visual system is complex and consists of several interacting anatomical structures. Here, we will describe the process of how photons of light from our surroundings become signals that the brain turns into representations of our surroundings.
Sensation vs. perception
The following 3 chapters are focused on sensation and perception. By definition, the words “sensation” and “perception” are very similar. However, in psychology and neuroscience, the two have slightly different meanings. Both terms are related to processes of getting information from the environment. “Sensation” is the detection of stimuli, whereas “perception” is the interpretation of those stimuli. The quantity of stimuli that you sense far outnumbers what can be realistically perceived. For example, imagine you are at a loud party, having a conversation with someone. You sense so many things here: all the friends and strangers you see, the smells of sweat or food, and the sounds of music and various voices. But while speaking to your friend, all you perceive is the person in front of you while all the other stuff fades into the background.
Perception is also shaped by past memories. Consider the following scenario. You are at home, studying by yourself quietly, when you hear footsteps and the jingling of keys. This process is sensation. Based on the fact that you have a roommate, and you know they have a nightly routine of studying late at the library before returning home, you interpret those sounds as your roommate coming home. This process is perception.

Chapter 7 outline
7.1 The Eye
7.2 The Retina
7.3 The Optic Nerve
7.4 Visual Perception in the Brain
7.1 The Eye
Visual sensation starts at the level of the eye. The eye is an organ that has evolved to capture photons, the elementary particle of light. Photons are unusual because they behave as both particles and as waves, but neuroscientists mostly focus on the wave-like properties. Because photons travel as waves, they oscillate at different frequencies. The frequency at which a photon oscillates is directly related to the color that we perceive. The human visual system is capable of seeing light in a very narrow range of frequencies. On the short end, 400 nm wavelengths are observed as violet, while on the long end, 700 nm wavelengths are red. Ultraviolet light oscillates at a wavelength shorter than 400 nm, while infrared light oscillates at a wavelength longer than 700 nm. Neither ultraviolet nor infrared light can be detected with our eyes.
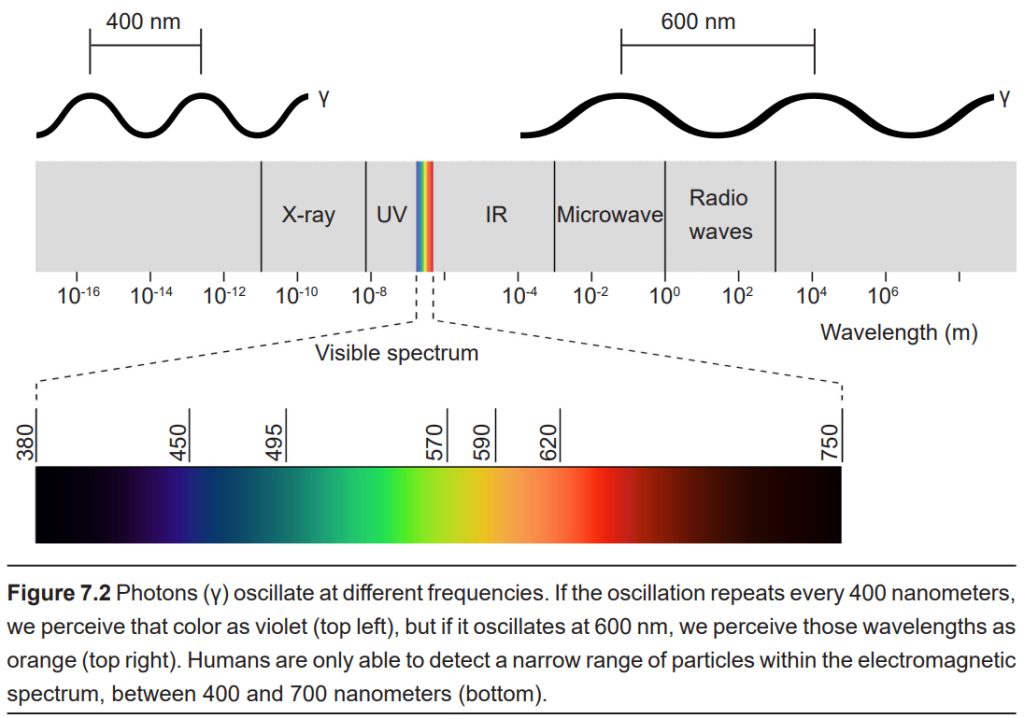
For us to see light, three basic conditions need to be met. First photons need to be present. When there are no photons of light, as in the depths of the Mammoth caves, we are unable to detect anything.
Secondly, photons of light must reflect off objects in the surrounding world. If photons cannot bounce off the object, the eye cannot collect the photons. Black appears colorless since it absorbs more photons, while pure white reflects photons. The wavelengths that reflect off objects determine the color that we perceive.
Third, photons need to reach the back of the eyeball. Photons pass through several anatomical structures before the nervous system processes and interprets them.
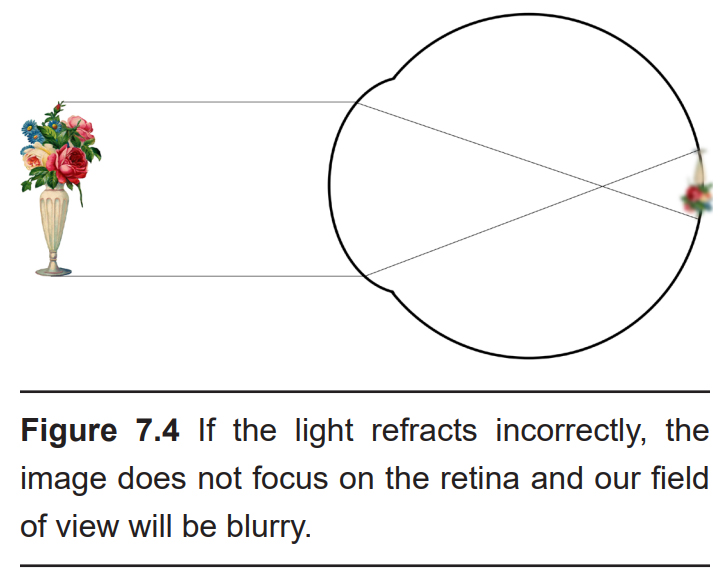
The anterior, or front-most part of the eye, is the cornea. The cornea refracts, or bends, the incoming rays of light so that they converge precisely at the retina, the posterior most part of the eye. If the light rays fail to properly converge, a person would be near-sighted or far-sighted, and this would result in blurry vision. Glasses or contact lenses bend light before it reaches the cornea to compensate the cornea’s shape.
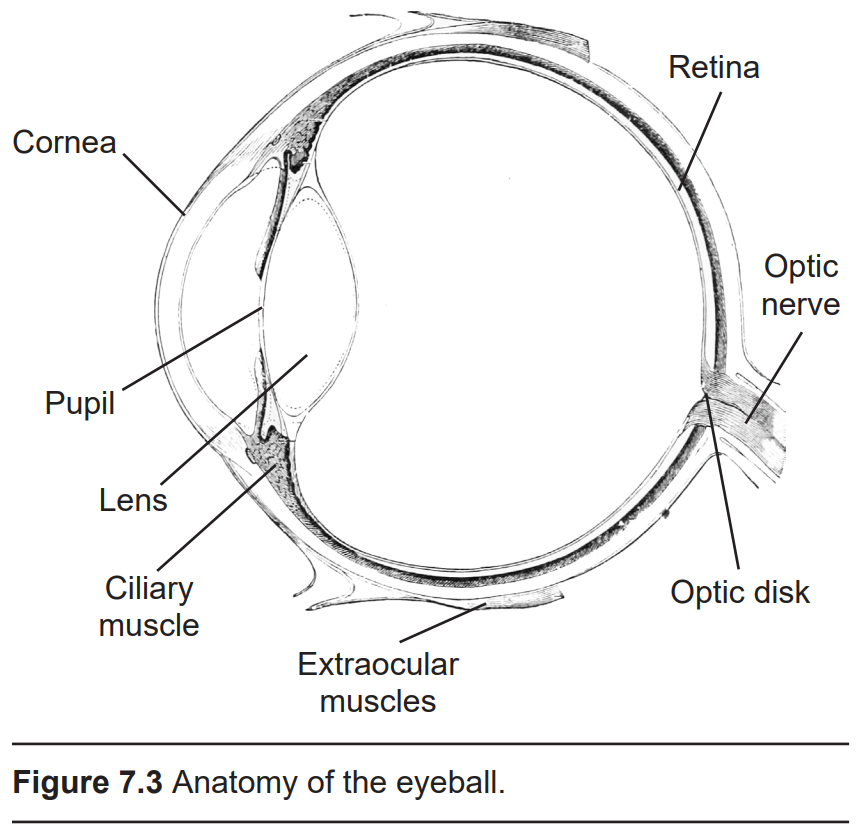
After passing through the cornea, light enters through a hole in the center of the eye called the pupil. The diameter of the pupil can change depending on ambient light conditions. In the dark, the pupil dilates (also called mydriasis), or gets bigger, which allows the eye to capture more light. In bright conditions, the pupil constricts (also called miosis), or gets smaller, which decreases the amount of light that enters the eye. Two muscles, the pupillary dilator muscle and the pupillary sphincter muscle, are responsible for change in pupil size. The pupillary dilator muscle causes dilation and is downstream of the sympathetic branch of the autonomic nervous system, while the pupillary sphincter causes constriction and is activated by the parasympathetic nervous system.
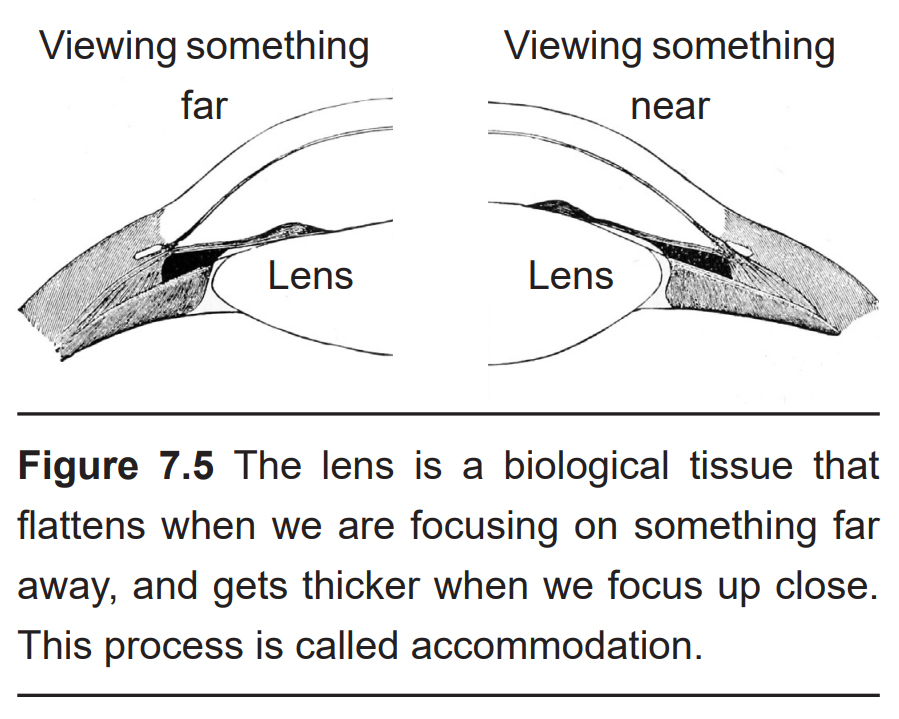
The next structure that light passes through is the lens. Like the cornea, the lens refracts light so that the rays converge on the back of the eye. The lens of the eye is shaped like a camera lens or a magnifying lens, convex on both sides (biconvex). Unlike the cornea, the lens is capable of adjusting its shape, a process called accommodation. A circular muscle that surrounds the lens, called the ciliary muscle, changes the shape of the lens depending on the distance of the object of focus. Whenever you focus your eyes on something close, such as holding your hand a few inches away from our face, the lens becomes thicker in the middle. Conversely, when you focus on the distant horizon, the lens flattens.
Another interesting consequence of the shape of the lens is the way it bends light. Think back to when you’ve held a magnifying lens in front of you as you look through it: The world you see through the lens is both upside down and reversed. This is exactly what the lens in our eyes does as it refracts light. As we look straight ahead, light from the upper part of our vision, like the ceiling, gets projected to the bottom of the eyeball, while light from the floor passes through the lens and refracts to the upper part of the eyeball. Additionally, light from the left part of our environment is refracted onto the right part of the eye, and vice versa.
We use the term visual field to describe the portion of our surroundings that can be seen without moving your eyes. If you look straight ahead, information on the left half of our vision are in our left visual field, which is projected onto the right half of both eyes. Conversely, objects in the right half of our vision are in our right visual field, which is projected as images onto the left half of both eyes.
There are six muscles that control eye movement. These muscles, plus the muscle that controls upper eyelid movement, are called the extraocular muscles. These seven muscles receive innervation from three cranial nerves, specifically cranial nerves 3, 4, and 6.
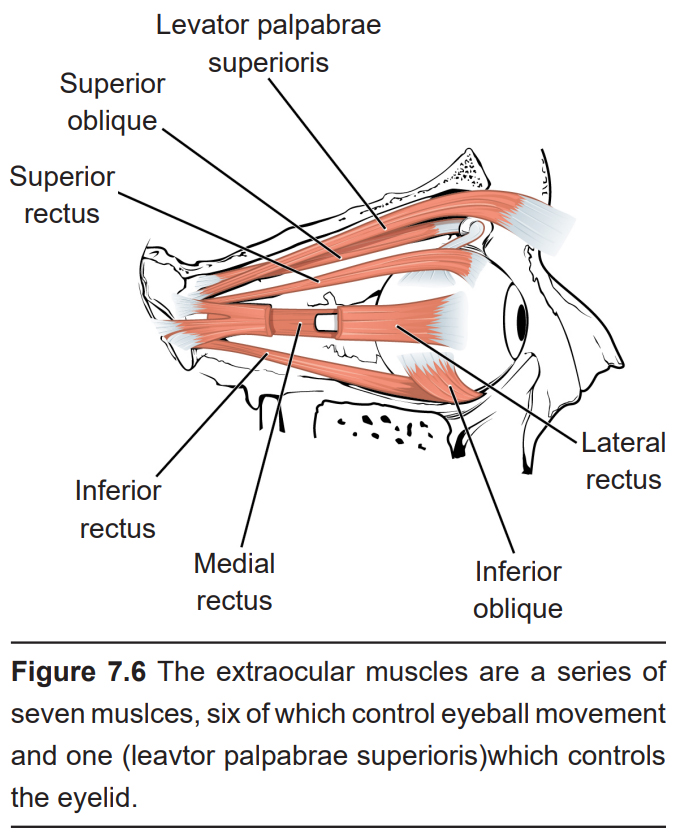
7.2 The Retina
The back of the eyeball is the retina (ret-n-UH). The medial half of the retina is also referred to as the nasal hemiretina since it is closer to the nose, while the lateral half of the retina is called the temporal hemiretina since it is closer to the temporal bone of the skull. The terms “upper hemiretina” and “lower hemiretina” are commonly used as well.
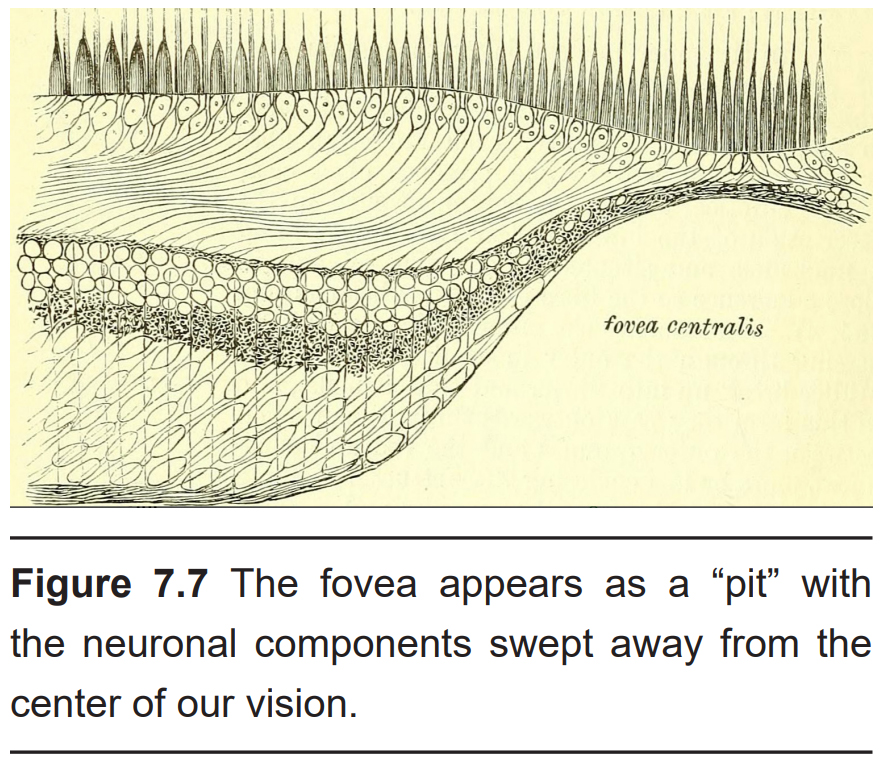
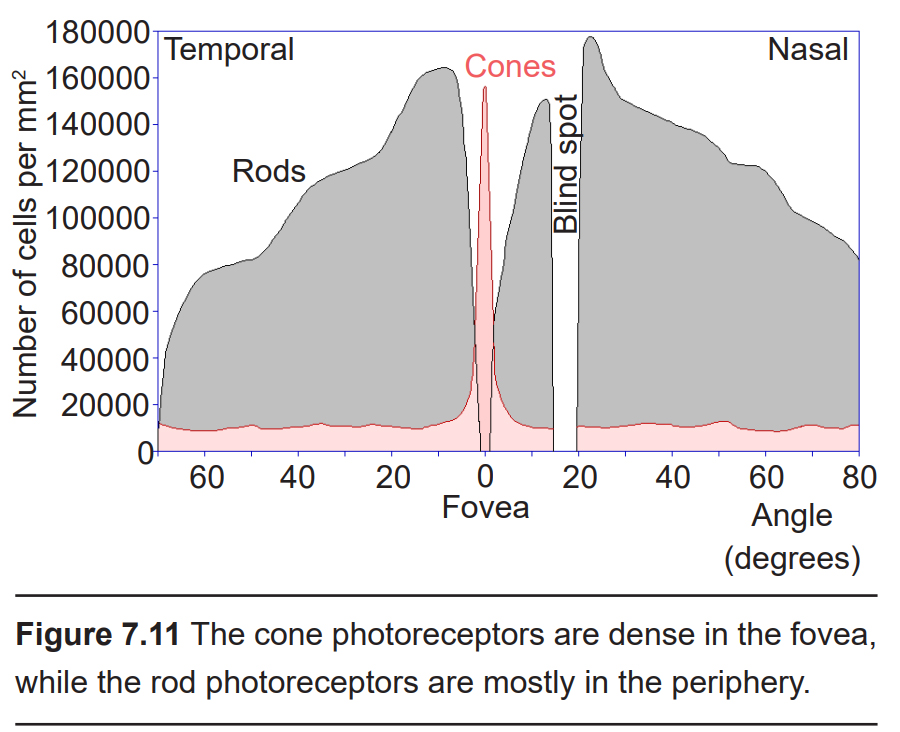
In the center of the retina is a small pit called the fovea. Light from only about 2 degrees from the center of the visual field gets focused onto the fovea. Compared to the rest of the retina, the fovea is where we have the highest acuity, or the clearest vision. As you are reading this text, light information from only 4 or 5 letters lands on the fovea.
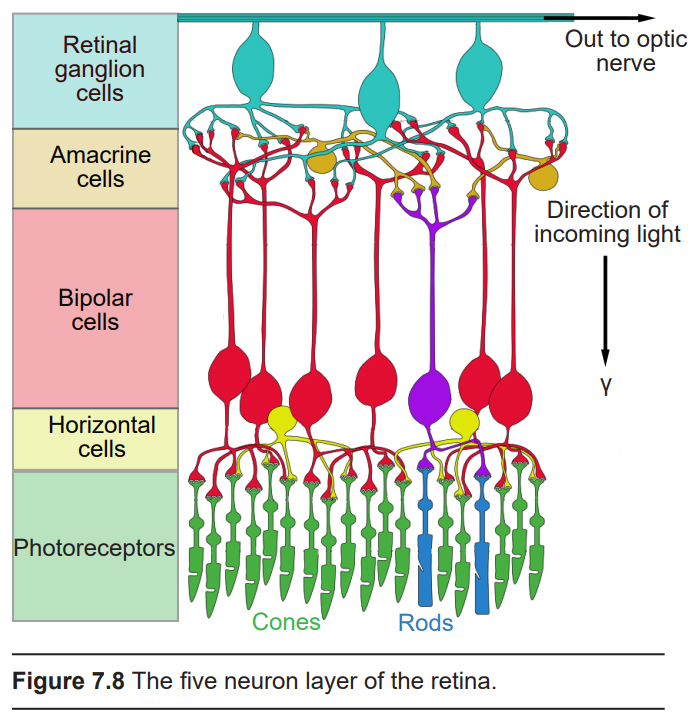
The retina is the beginning of the nervous system’s involvement with the visual system. It consists of five layers of neurons (described below). The pathway of information moves from the back of the eye towards the front, before exiting the back of the eye in an area called the optic disk. Visual information is passed directly through three of the five neuronal layers before being shuttled into the brain via the optic nerve. The other two layers of cells modify that route of communication. In the next paragraphs, we’ll take an in-depth look at the role of each of the five layers of neurons in the retina.
In the order of the pathway of communication, the five neurons of the retina are:
7.2.1 Photoreceptor cells
Photoreceptors are the first cells in the neuronal visual perception pathway. They are the cells that detect photons of light and convert them into neurotransmitter release, a process called phototransduction.
Morphologically, photoreceptor cells have two parts, an outer segment and inner segment. The outer segment contains stacks of membranous disks bounded within the neuronal membrane. These membranous disks contain molecules called photopigments, which are the light-sensing components of the photoreceptors. Hundreds of billions of these photopigments can be found in a single photoreceptor cell.
The inner segment contains the nucleus and other organelles. Extending from the inner segment is the axon terminal. Photoreceptors are classified into two categories, named because of their appearance and shape: rods and cones.
Rod photoreceptor cells
Visual information from our peripheral vision is generally detected by our rod cells, which are most densely concentrated outside the fovea.
Rod cells are organized to have high synaptic convergence, where several rod cells (up to 30) feed into a single downstream route of communication (the bipolar cells, to be specific – see 7.2.3). An advantage of a high-convergence network is the ability to add many small signals together to create a seemingly larger signal. Consider stargazing at night, for example. Each rod is able to detect low levels of light, but signals from multiple rod cells, when summed together, allows you to recognize faint light sources as a star. A disadvantage of this type of organization is that it is difficult to identify exactly which photoreceptor is activated by the incoming light, which is why accuracy is poor when seeing stimuli in our peripheral vision. This is one of the reasons that we cannot actually read text in our peripheral vision or see the distinct edges of a star.
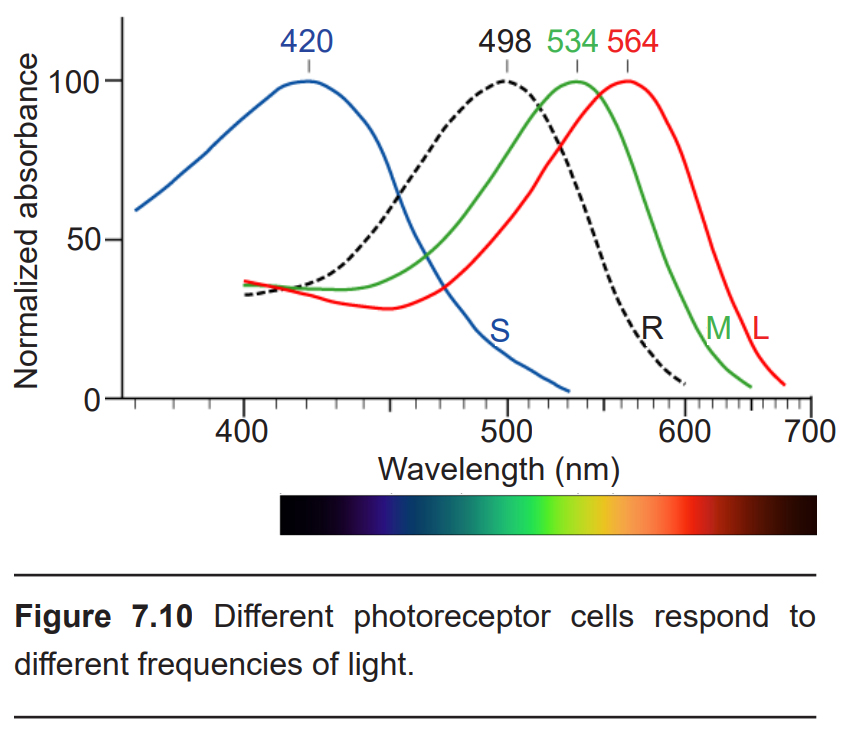
Rod cells are most sensitive to light that has a wavelength of 500 nm (blue-ish green). Light at other wavelengths still causes changes in the way rod cells respond, but to a lesser degree. Rod photoreceptors are maximally active in low-light conditions, which is why our surroundings appear to have a blue-ish tint at night. This phenomenon is known as the Purkinje shift, named after the Czech anatomist Jan Purkinje who observed that his favorite brightly- colored flowers appeared dull and more blue as the sun was setting.

Cone photoreceptor cells
Cone photoreceptor cells allow for high-acuity vision. They are most densely packed at the fovea, corresponding to the very center of your visual field. Despite being the cell population that we use for our best vision, cone cells make up the minority of photoreceptors in the human retina, outnumbered by about 20-times more rod cells.
Unlike rod cells, cone cells have very low synaptic convergence. In fact, at the point of highest visual acuity, a single cone photoreceptor communicates with a single pathway to the brain. The signaling from low-convergence networks is not additive, so they are less effective at low light conditions. However, because of this low-convergence organization, cone cells are highly effective at precisely identifying the location of incoming light.
Cone photoreceptors are responsible for processing our sensation of color (the easiest way to remember this is cones = color). The typical human has three different types of cone photoreceptors cells, with each of these three types tuned to specific wavelengths of light. The short wavelength cones (S-cones) respond most robustly to 420 nm violet light. The middle wavelength cones (M-cones) exhibit peak responding at 530 nm green light, and the long wavelength cones (L-cones) are most responsive in 560 nm red light. Each of these cones is activated by other wavelengths of light too, but to a lesser degree. Every color on the visible spectrum is represented by some combination of activity of these three cone photoreceptors.
The idea that we have two different cellular populations and circuits that are used in visual perception is called the duplicity theory of vision, and is our current understanding of how the visual system perceives light. It suggests that both the rods and cones are used simultaneously and complement each other. One component, called the photopic vision system, uses cone photoreceptors of the retina, and is responsible for high-acuity sight and color vision in daytime. Its counterpart, called scotopic vision, uses rod photoreceptors and is best for seeing in low-light conditions, such as at night.
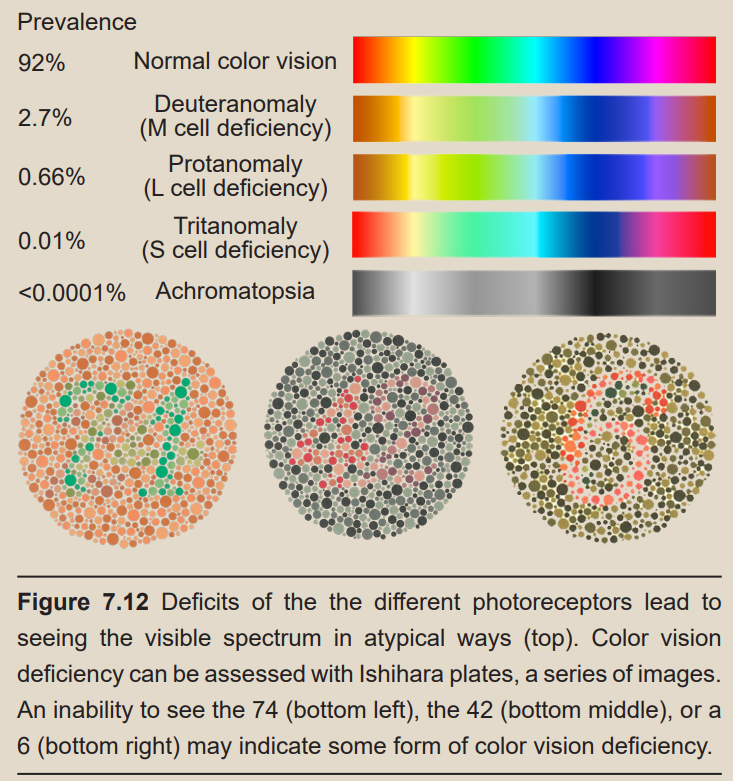
Clinical connection: Color vision deficiency Color vision deficiency is a very common condition resulting from a dysfunction in cone photoreceptor cells. As you know, each person has three types of cone cells. However, if one of the three types of cone cells fails to respond to the correct wavelengths of light, the person will lose the ability to differentiate between certain colors on the visible light spectrum. For example, lacking functional L-cones causes a person to have difficulty distinguishing purple from blue.
Color vision deficiency can either be acquired (as a symptom of disease or as a side effect of exposure to certain chemicals) or inherited. Most cases are inherited, and the genes associated with color vision deficiency are often recessive and located on the X-chromosome. Because these genes are sex-linked, color vision deficiency is 15 times more likely to occur in men than in women with a prevalence of about 8% of males and less than 1% in females.
On the molecular and cellular scale, the neuronal encoding of light information is counterintuitive. We normally think of neurons as being excited when they are activated, releasing more neurotransmitter upon experiencing a stimulus. Photoreceptors, on the contrary, decrease in excitability when they are exposed to light. Additionally, they do not fire action potentials. Instead, their membrane potential fluctuates, which determines the amount of glutamate release: depolarization causes more glutamate release, and hyperpolarization causes a decrease in glutamate release.
In the dark, photoreceptor cells have leak sodium channels that allow for extracellular sodium to enter those neurons, which is called the dark current. This current causes cellular depolarization. When a photon of light strikes the photoreceptor, the cell responds by decreasing its intracellular stores of a signaling molecule called cGMP, which in turn decreases the dark current. Light causes the neuron to become hyperpolarized, which inhibits neurotransmitter release.
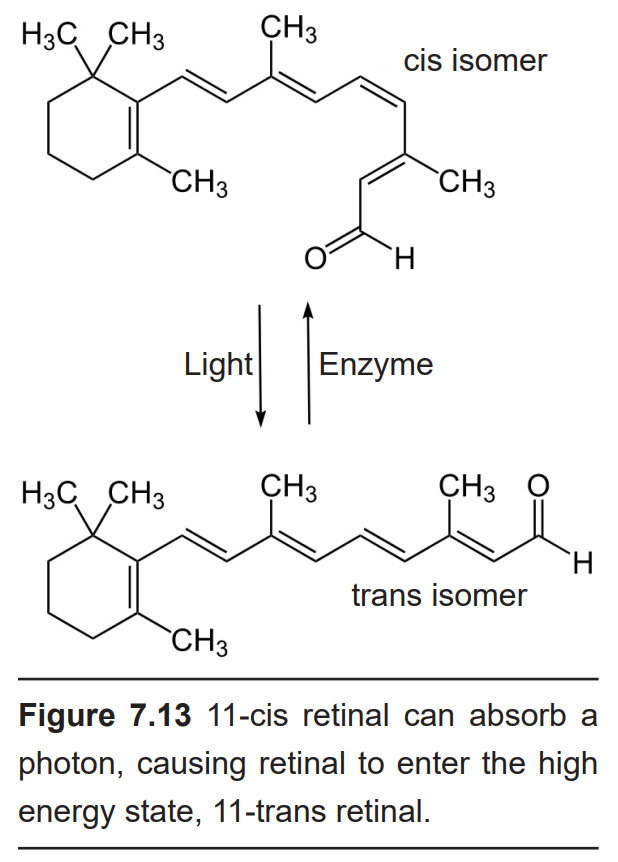
These neurons sense light because of the presence of the photopigments found in the outer section of the photoreceptors. The photopigment molecule has two components. Retinal is a chemical synthesized from Vitamin A. In the dark, retinal exists as 11-cis- retinal, which has a low energy molecular configuration. Light can be highly energetic. When a photon hits retinal, it causes the chemical to change to all- trans-retinal, which is the high energy state configuration. While in the all-trans- retinal configuration, cGMP levels are decreased, therefore blocking the dark current, which causes hyperpolarization.
The other component is a protein called an opsin. Opsins exist in different forms, and they cause retinal to respond differently depending on the wavelength of light: some opsins cause the photoreceptors to be maximally activated at 420 nm, others at 530 nm, and so on.
Sidebar: Lateral inhibition
Bring your vision over to one of the white “intersections” in the Hermann grid below. In your peripheral vision, do you see faint fuzzy outlines of gray circles at the other white intersections?
This phenomenon happens because of the circuitry of the horizontal cells. In being activated, they block adjacent signaling pathways from also sending information forward. Because horizontal cells inhibit adjacent neurons at the same “level,” we describe this phenomenon as lateral inhibition. It is hypothesized that lateral inhibition evolved to allow for better edge detection, the ability to easily differentiate the outlines of objects.
Now bring your focus to one of those fuzzy gray blobs. You will notice that the grayness disappears immediately when we look directly at it, but a new gray blob appears where you were just looking at. This happens because the size of the receptive field for each photoreceptor-to-bipolar synapse differs at the fovea from the periphery. Foveal photoreceptors have very little convergence onto their bipolar cells, so they have very high precision, and are therefore less susceptible to the illusion as seen above.

7.2.2 Horizontal cells
Horizontal cells are interneurons that do not directly participate in the pathway of signaling in visual perception. Rather, they communicate laterally, receiving synaptic inputs from photoreceptors, and forming synaptic connections with the axons of nearby photoreceptors. Like most interneurons in the nervous system, horizontal cells are inhibitory.
Photoreceptors release glutamate onto the horizontal cells, which causes the horizontal cells to depolarize. More glutamate is released when the photoreceptors are in the dark. In the light, the amount of glutamate released is decreased, which causes horizontal cells to hyperpolarize. This hyperpolarization then decreases neurotransmitter release onto nearby photoreceptor axons. Less inhibition onto these photoreceptors causes depolarization, which decreases signaling from these adjacent photoreceptors.
This complex pattern of neurotransmission can be summarized more simply: activation of a single photoreceptor causes inhibition of adjacent photoreceptors. The activity of horizontal cells helps us isolate the origin of a light signal by minimizing the noise around it.
7.2.3 Bipolar cells
Bipolar cells are the second type of neuron that is directly responsible for visual perception, receiving visual input signals from photoreceptor cells. They are named because of their morphology: from the cell body, they extend projections in two opposite directions. Their dendrites receive neurotransmitter signals from photoreceptors, and they send axonal projections to the next two layers of neurons in the retina (amacrine cells and retinal ganglion cells).
In mammals, bipolar neurons receive inputs exclusively from either rod or cone photoreceptors. In other words, there are no bipolar neurons that receive both rod and cone photoreceptor input.
As we’ve already discussed, photoreceptor cells release glutamate. At most synapses in the nervous system, glutamate is excitatory. However, at synapses between photoreceptors and bipolar cells, glutamate activates metabotropic mGluR6 receptors, which are inhibitory. In the light, there is a decrease in glutamate release from the photoreceptors, and this leads to an increase in bipolar cell activity.
7.2.4 Amacrine cells
Amacrine cells, like the horizontal cells, are interneurons that modify the communication pathway rather than directly being involved with light sensation. Amacrine cells receive inputs from bipolar cells, and in turn, modulate the activity of nearby neurons. Like most interneurons, amacrine cells synthesize and release the inhibitory neurotransmitter GABA.
Amacrine cells exist in several different shapes and sizes. They help us detect directional motion, aiding us in knowing when to transition between our scotopic or photopic vision systems. Some amacrine cells also perform circadian regulatory functions.
7.2.5 Retinal ganglion cells
Retinal ganglion cells are the third and last cell type that directly conveys visual sensory information, receiving inputs from the bipolar cells. The axons of the retinal ganglion cells bundle together and form the optic nerve, which then exits the eyeball through the optic disk. These axons are unmyelinated before leaving the optic disk – if they were myelinated, the lipids that make up the myelin sheath would diffract light and cause imperfections in vision.
Unlike the two cells before them (the photoreceptors and bipolar cells), retinal ganglion cells communicate using action potentials.
Although most retinal ganglion cells receive light information through communication downstream of the photoreceptors and bipolar cells, a small population of retinal ganglion cells (1-2%) are photosensitive. These cells contain a molecule called melanopsin, a light- sensitive opsin molecule just like the photopsin or rhodopsin found in photoreceptors. These cells contribute significantly to our body’s ability to adjust the circadian rhythms based on external light cues (see chapter 12 for more information.)
Sidebar: The blind spot
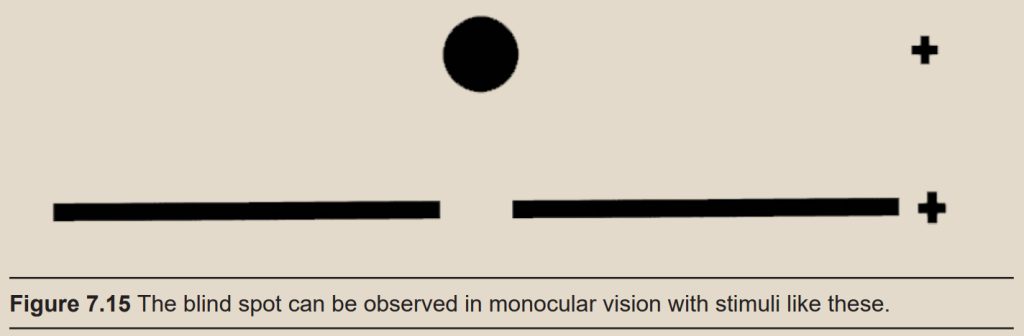
Take a look at the image below. Close your right eye and bring your face close to the page, focusing on the plus sign. Slowly move your head away from the page. At a distance of about one and a half feet away, the dot will disappear!
The dot disappears under these specific conditions because visual information from the dot falls directly onto the blind spot. The blind spot is about 15 degrees temporal from the fovea. We don’t notice the blind spot in everyday life because usually both eyes are working simultaneously, so when something enters the blind spot of the right eye, it appears in the visual field of the left eye. The blind spot only comes into effect when using monocular vision, or when we only see with one eye. The blind spot is elliptical, and you can demonstrate this using the drawing above: Bring the dot into your blind spot as before, but this time, rotate the page, pivoting around the plus sign. The dot reappears as it moves above or below the blind spot.
Now, repeat with the figure below. Hold the page at the same distance where you noticed the blind spot. How would you describe what happens to the break in the line? While we are unable to physically detect any light that falls onto the blind spot, our brains fill in the gap in sensation by “filling in” the blind spot with the adjacent visual stimuli.
The retina is not completely uniform across the entire back of the eye. There are a few spots of particular interest along the retina where the cellular morphology is different: the fovea and the optic disk.
There are two cellular differences that explain why the fovea is the site of our best visual acuity. For one, the neurons found at the fovea are “swept” away from the center, which explains why the fovea looks like a pit. Cell membranes are made up mostly of lipids, which distort the passage of light. Because there are fewer cell bodies present here, the photons of light that reach the fovea are not refracted by the presence of other neurons.
Secondly, the distribution of photoreceptors at the fovea heavily leans toward cone type photoreceptors. Because the cone cells at the fovea exhibit no convergence, they are most accurately able to pinpoint the exact location of incoming light. On the other hand, most of the photoreceptors in the periphery are rod cells. With their high-convergence circuitry, the periphery of the retina is suited for detecting small amounts of light, though location and detail information is reduced.
Another anatomically interesting area of the retina is an elliptical spot (1.9mm vertical by 1.75 mm horizontal) called the optic disk. This is where the optic nerve exits the eye. At this part of the retina, there is an absence of photoreceptor cells. Because of this, we are unable to perceive light that falls onto the optic disk. This spot in our vision is called the blind spot.
7.3 The Optic Nerve
The optic nerve, or cranial nerve II, exits the posterior end of the eyeball, and travels posteriorly along the ventral surface of the brain. Like all other cranial nerves, the optic nerve is paired, meaning there is one on each half of the body – one for each eye. Both optic nerves merge at a spot called the optic chiasm, then diverge yet again as they travel posteriorly towards the thalamus.
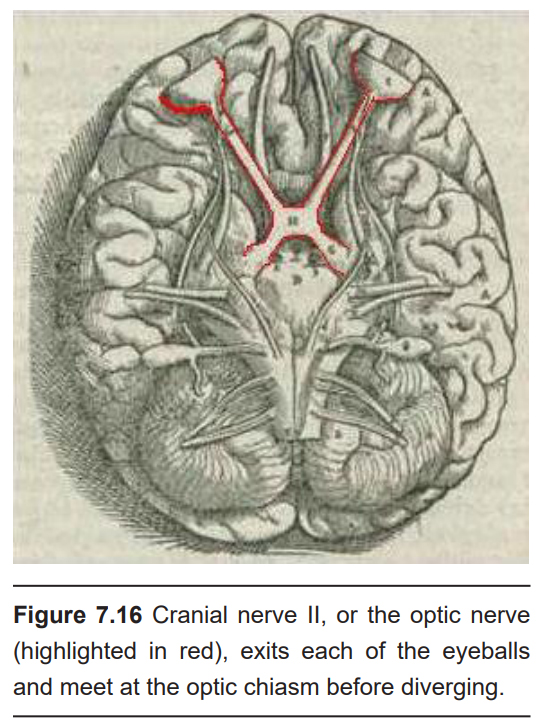
The axonal connections in the optic nerve are not quite as simple as its anatomical appearance. From each optic nerve, some of the nerve fibers cross the midline, headed towards the contralateral hemisphere. Other nerve fibers meet at the optic chiasm, but then project into the ipsilateral hemisphere. Visual information from the nasal hemiretina projects contralaterally, so the left nasal hemiretina crosses at the optic chiasm to project into the right hemisphere of the brain. On the other hand, visual information from the temporal hemiretina projects ipsilaterally (left temporal hemiretina projects into left hemisphere, and right temporal hemiretina projects into right hemisphere ).
The easy way to keep track of this unusual system is to remember that all information from the left visual field enters the right hemisphere of the brain, while visual information from the right visual field enters the left hemisphere of the brain. Because of the light-refracting properties of the lens, objects in the left visual field get represented onto the right half of each eyeball.
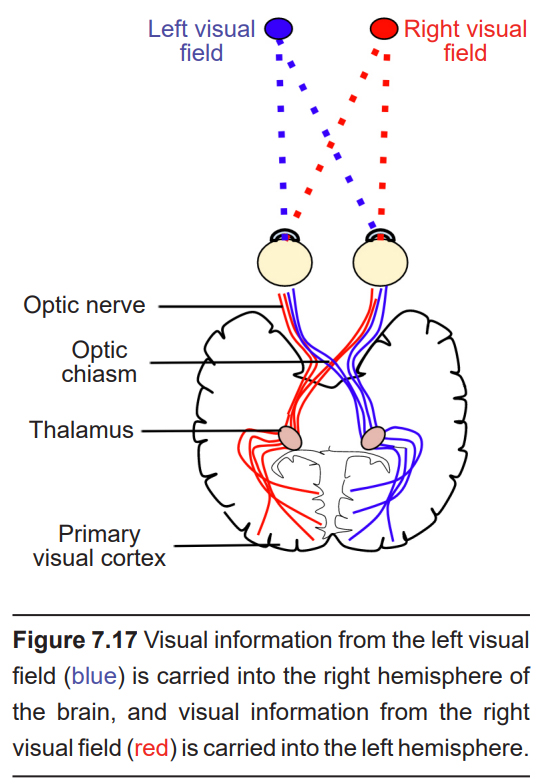
The right half of the left eye is the nasal side, so this information crosses contralaterally. The right half of the right eye is the temporal side, so this stays ipsilateral. This information then passes towards the thalamus for the beginning of central processing.
Not all of the axons convey direct visual information into the thalamus for visual perception. One pathway of the optic nerve sends information into a midbrain structure called the pretectal area. The information sent into the pretectum is related to some of the more “primitive” visual functions. For example, the ability for the eyes to follow something unconsciously as it moves across your field of view is called smooth pursuit, and this reflex is partly carried out by the optic nerve pathway into the pretectal area.
Another signaling pathway that branches from the optic nerve is the retinohypothalamic tract (RHT). It does not carry any conscious visual information. The RHT conducts light information from the intrinsically-photosensitive retinal ganglion cells. Instead of sending axonal projections towards the thalamus, this information pathway forms a synapse into a region of the hypothalamus called the suprachiasmatic nucleus, or SCN. This structure functions to help the body adapt its sleep-wake cycle in the face of changing day- night patterns (Chapter 12).
Clinical correlation: Glaucoma
The optic nerve is particularly sensitive to injury. A condition called glaucoma can cause optic nerve destruction, leading to blindness. Glaucoma affects up to 3 million Americans, but can be easily detected during an annual eye exam. Glaucoma can result from several different causes including high intraocular pressure, physical trauma, medications, or a natural genetic weakness of the nerve. High intraocular pressure is the greatest risk factor for glaucoma, since hypertension compresses the axons, which can also decrease the blood flow to the optic nerve. Almost all the first-line therapies to treat glaucoma decrease intraocular pressure.
7.4 Visual perception in the brain
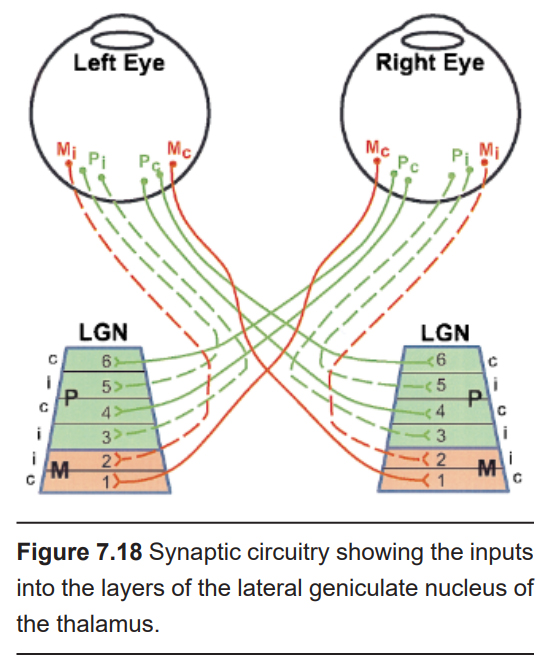
The first synapse of the optic nerve is formed in the thalamus at a subregion called the lateral geniculate nucleus, or LGN (originating from the Latin genu-, meaning “knee”). The LGN is divided into six layers, named 1 through 6, going from ventral to dorsal. Histologically, the LGN is made up of three different populations of neurons. The magnocellular cells (magn– means large, as in “magnify” or “magnanimous”), or M cells, are large-diameter neurons (~20 µM) that are found in layers 1 and 2. The parvocellular cells (parv– means small), or P cells, are small- diameter (~10 µm) neurons found in layers 3 through 6. Ventral to each layer is a row of very small neurons, called koniocellular cells (konio– means dust), or K cells.
We know that the visual information from the temporal retina synapses ipsilaterally, while information from the nasal retina synapses contralaterally. The specific synaptic inputs into the LGN follow a specific organization depending on the origin of the retinal ganglion neurons. Temporal retinal ganglion neurons synapse only onto layers 2, 3, and 5 of the ipsilateral LGN, while nasal retinal ganglion neurons form synapses exclusively onto contralateral layers 1, 4, and 6.
The outputs of the LGN are a series of axonal bundles called the optic radiations. These optic radiations are divided into two main bundles, the upper and lower divisions. The upper divisions carry information from the lower visual field, whereas the lower divisions carry information from the upper visual field.
From the LGN of the thalamus, the optic radiations project to the occipital lobe at the caudal (posterior) end of the brain. Once visual information travels into the cortex, the process is less about sensation and mostly about perception. For example, many of the post-optic radiation signaling pathways are concerned with questions like attention, object orientation, and recognition. Many visual perception functions are performed by multiple brain regions, and no single area is responsible for any one compartmentalized function. In other words, there is still much work to be done on understanding the precise function of the areas of the visual cortex.
The outputs of the LGN are axons which form synapses in the primary visual cortex, which is also called V1. Alternatively, V1 is also called the striate cortex because it has a large white stripe that can be seen in surgical dissection. This white stripe is the bundle of incoming optic tract axons, which are heavily myelinated.
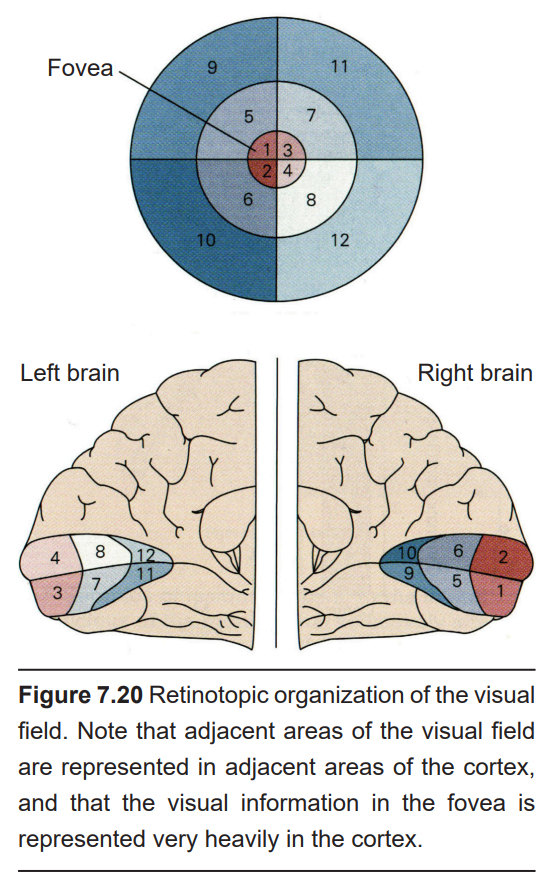
Each neuron in V1receives visual information from a specific patch of retinal cells. This organizational pattern, where a section of retinal inputs map onto neurons of a specific section of V1, is called retinotopic organization. For example, the dorsal part of V1 receives information from the bottom half of the visual field. Visual information from the fovea, despite being only 1% of the total visual field, takes up about half of all neurons in V1.
After processing in V1, visual information is passed along to the secondary visual cortex, or V2. V2 represents the beginning of the visual association cortices. Following V2 are other visual cortical areas that contribute to various aspects of visual perception. For example, V3 and V5 help us comprehend motion, V4 contributes to color perception, and V6 helps with understanding our position in our surroundings.
Clinical connection: Anatomy of vision loss
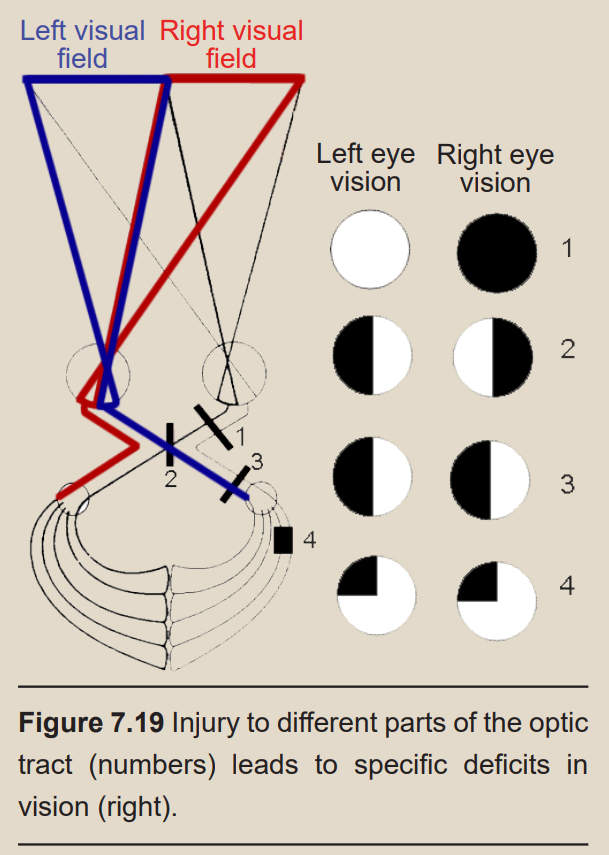
Health care professionals can diagnose the location of an injury based on the deficits in a patient’s visual field. To record the sites of vision loss, they may use a pair of circles which show which areas of the visual field are lost. For example, if a person is missing their right eye, all information from the right eye will be missing and the right circle will be colored in black.
If the visual information-conveying pathway is damaged, a person may suddenly lose half of their visual field resulting in a condition called hemianopsia (hemi– meaning half, a– meaning none, and –opsia referring to vision). There are a few forms of hemianopsia that correlate to specific injuries of the optic nerve. For example, injury to one branch of the nerve after the optic chiasm results in homonymous hemianopsia, the loss of either the left or right visual field of both eyes.
Injuries may damage some optic radiations while sparing the others. This results in a condition called quadrantanopsia, resulting in a loss of a fourth, or one quadrant, of a person’s visual field. If the upper division of the right optic radiation is injured, the person will lose the upper left visual field of both of their eyes: left superior quadrantanopsia.
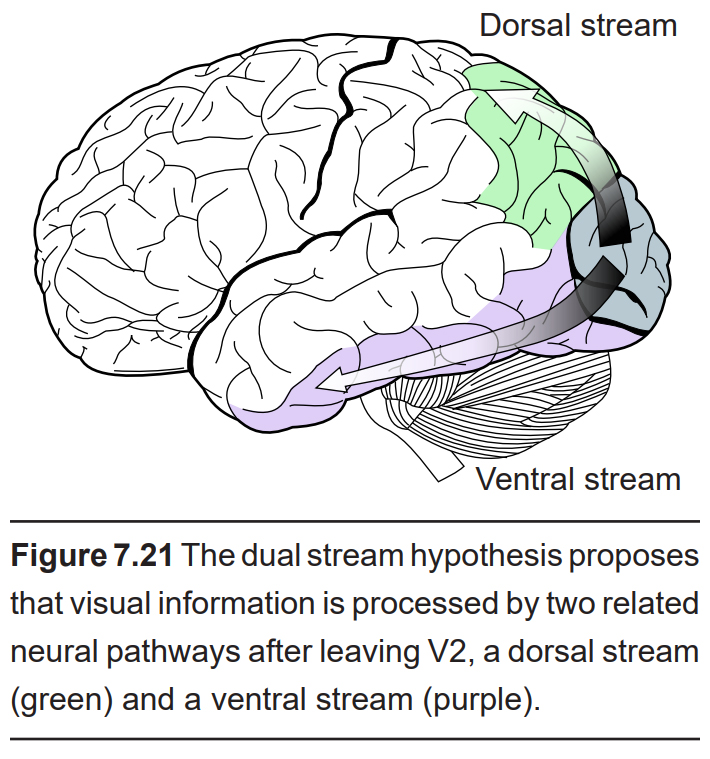
Dual stream hypothesis
After V2, visual information passes through two streams of communication: the dorsal stream and the ventral stream.
The dorsal stream is described as the “where” pathway because these structures help us identify where objects are located in the space around us. The dorsal stream includes area V5, which contributes to perception of motion. These structures also guide us when we move through our environments, contributing to our sense of spatial awareness. For example, a task such as reaching out to grab an object in front of you uses a combination of these features, so this task is guided largely by dorsal stream structures.
The ventral stream is the “what” pathway, and helps in the identification of objects that we see. Ventral stream structures are important for visual memory. The fusiform face area (FFA) and the parahippocampal place area (PPA) are especially important for the storage of facial stimuli and scenery or buildings, respectively. When the visual system senses these complex stimuli, those signals get processed through these ventral stream pathways. These incoming stimuli are compared with the memories stored in the ventral stream, and this comparison contributes to our capacity for perception and identification.
The two streams are not independent of each other. Rather, successful organisms require the melding of both components of visual perception. For example, imagine you are a prehistoric organism living in a food-scarce environment. Approaching a small berry tree, you would use ventral stream structures to correlate the berries with memories: Did these berries taste good and give me the calories I need to survive? Or did these berries make me violently ill, and are therefore probably poisonous? If they are the delicious berries that I want, I will use the dorsal stream structures to take note of their precise location so I can reach out for them and pick the berries . In this example, proper interaction of the dual streams contributes to goal-driven actions.
Clinical connection: Dysfunction of the dual-stream
Deficits in the circuitry of visual processing up until this point in the chapter result in largely predictable outcomes. However, once that information enters the cortex of brain, the circuitry involved becomes much more complex, and unusual visual perception disorders may result from atypical neural communication in the brain.
Akinetopsia, or motion blindness, is a very rare visual condition resulting in the inability to perceive objects in motion. Instead of seeing movement, these people see the world as a series of “freeze frames” that refresh occasionally, or only whenever the object stops moving. People with akinetopsia have severe difficulty crossing the street or pouring a cup of tea. Usually, akinetopsia results from brain injury like trauma or a stroke.
Prosopagnosia, or face blindness, is another unusual visual condition that results in people who have tremendous difficulty with identifying faces. People with prosopagnosia recognize that they are looking at a face. They see the eyes, nose, and mouth, but they often cannot identify who that face belongs to. Some cases are so severe that they cannot even recognize themselves in a mirror. Prosopagnosia is far more common than akinetopsia; an estimated 1% of people have some degree of face blindness.
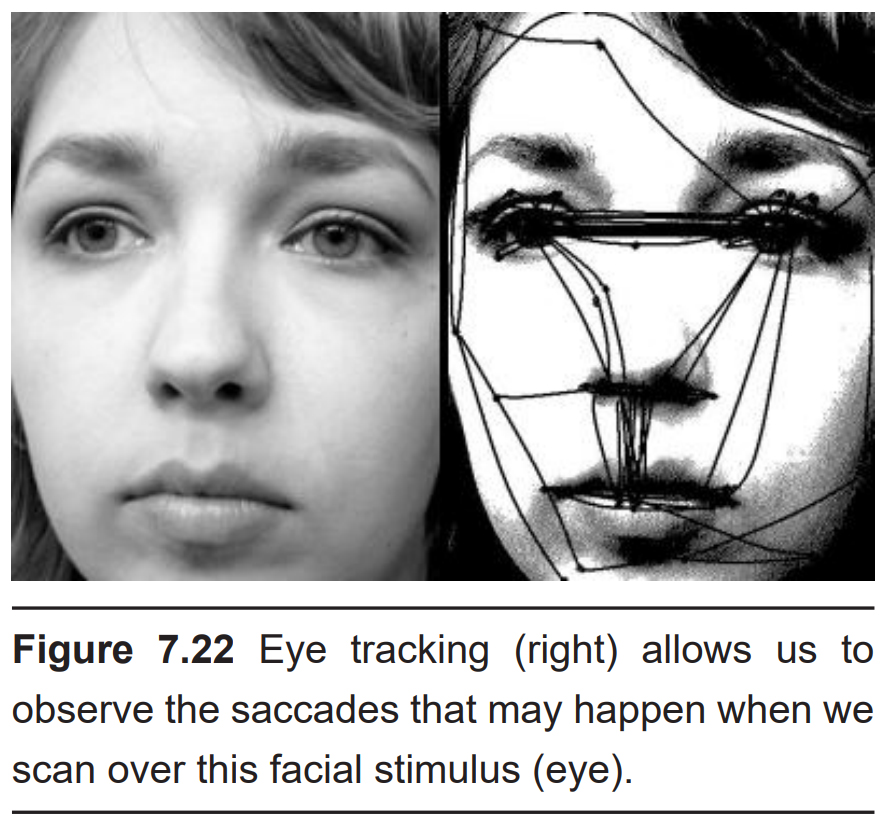
Saccadic eye movements
Take a look at the objects around you right now. You don’t notice it, but as you shifted your focus from one object to the next, your eyes made tiny, rapid, jerking motions over the different objects instead of moving smoothly across your field of vision. These rapid movements, driven by the actions of the extraocular muscles, are called saccades (pronounced “suh-KAHD”). Saccades are among the fastest muscle movements in the body.
These saccades help you accurately capture visual information from your surroundings. The fovea, which has the highest acuity vision, only collects information from a tiny portion of your visual field. By performing these saccades, you can scan over several objects with the highest acuity, thus allowing your brain to build a representation of the visual field.
The Open Neuroscience Initiative is funded by a grant from the Vincentian Endowment Fund of DePaul University.
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License.
Image Credits
Cover: Niagara66 / CC BY-SA (https://creativecommons.org/licenses/by-sa/4.0) https://upload.wikimedia.org/wikipedia/ commons/1/19/Mammoth_Cave_National_Park_01.jpg
7.1 Image by Anja from Pixabay (https://pixabay.com/photos/key-keychain-door-key-house-keys-453480/ ) Image by Ann San from Pixabay (https://pixabay.com/photos/key-door-opening-door-lock-2732312/ )
7.2 https://upload.wikimedia.org/wikipedia/commons/3/30/EM_spectrumrevised.png modified by Austin Lim
7.3 https://upload.wikimedia.org/wikipedia/commons/d/dd/The_outlines_of_anatomy%2C_physiology%2C_and_hygiene._Being_an_edition_of_The_essentials_of_anatomy%2C_physiology%2C_and_hygiene%2C_rev._to_conform_to_the_legislation_making_the_effects_of_alcohol_and_other_%2814596873080%29.jpg
7.4 Image by Karen Arnold from Pixabay (https://pixabay.com/illustrations/vintage-flowers-vase-1772583/ ) 7.5https://upload.wikimedia.org/wikipedia/commons/0/06/PSM_V45_D217_Section_of_the_lens_showing_the_ mechanism_of_accommodation.jpg modified by Austin Lim
7.6 https://upload.wikimedia.org/wikipedia/commons/7/74/1107_The_Extrinsic_Eye_Muscles_right_eye_lat.png modified by Austin Lim
7.7 https://www.flickr.com/photos/internetarchivebookimages/14592617690/
7.8 https://upload.wikimedia.org/wikipedia/commons/e/e3/Rods_Cones_Synapse.svg Modified by Austin Lim
7.8 https://upload.wikimedia.org/wikipedia/commons/0/02/Red_geranium_Purkinje.jpg
7.10 https://commons.wikimedia.org/wiki/File:Cone-response-en.png#/media/File:Cone-response-en.png modified by Austin Lim
7.11 https://commons.wikimedia.org/wiki/File:Distribution_of_Cones_and_Rods_on_Human_Retina.png modified by Austin Lim
7.12 https://upload.wikimedia.org/wikipedia/commons/a/af/Color_blindness.png; https://upload.wikimedia.org/wikipedia/commons/e/e0/Ishihara_9.png; https://upload.wikimedia.org/wikipedia/commons/e/e0/Ishihara_9.png; https://commons.wikimedia.org/wiki/File:Ishihara_3.jpg
7.13 https://upload.wikimedia.org/wikipedia/commons/3/34/11-cis-Retinal_Vitamin_A-Aldehyd.svg
7.14 ttps://upload.wikimedia.org/wikipedia/commons/6/6f/Zludzenie_Hermana.png
7.16 https://upload.wikimedia.org/wikipedia/commons/5/5c/1543%2CVisalius%27OpticChiasma.jpg
7.17 https://upload.wikimedia.org/wikipedia/commons/4/4d/Neural_pathway_diagram.svg modified by Austin Lim
7.18 Reprinted with permission from Dr. Pasko Rakic (Early divergence of magnocellular and parvocellular functional subsystems in the embryonic primate visual system)
7.19 https://upload.wikimedia.org/wikipedia/commons/e/e5/Hemia.PNG modified by Austin Lim
7.20 https://commons.wikimedia.org/wiki/File:Retinotopic_organization.png modified by Austin Lim
7.21 https://commons.wikimedia.org/wiki/File:Ventral-dorsal_streams.svg modified by Austin Lim
7.22 https://upload.wikimedia.org/wikipedia/commons/d/d3/Szakkad.jpg
