5 Chapter 5: Signaling between Neurons
COVER IMAGE
Austin Lim, PhD (DePaul University)
Editor: Monica Javidnia, PhD (University of Rochester)
Previously, we described the electrical properties of a single neuron. A lone neuron can send action potentials as a means of communication, but cells become much more interesting when they have partners to talk to. The nervous system of the worm C. elegans is only 300 neurons, and yet it is complex enough to engage in moderately intricate behaviors like responding to repellant or attractant odors, social feeding, and long-term learning. The human brain, with its 86 billion neurons, can engage in these behaviors and so many more – only because of communication between the different neurons in the brain.
In this chapter, we will focus on the molecular-level features of communication between neurons, starting from the anatomical differences between synapses.
A lone neuron can send action potentials as a means of communication, but cells become much more interesting when they have partners to talk to. The nervous system of the worm C. elegans is only 300 neurons, and yet it is complex enough to engage in moderately intricate behaviors like responding to repellant or attractant odors, social feeding, and long-term learning. The human brain, with its 86 billion neurons, can engage in these behaviors and so many more – only because of communication between the different neurons in the brain.
In this chapter, we will focus on the molecular-level features of communication between neurons, starting from the anatomical differences between synapses.
Chapter 5 outline
5.1 Electrical vs. chemical synapses
5.2 Properties of vesicles
5.3 Receptors
5.4 Neurotransmitters
5.1 Electrical vs. chemical synapses
The synapse is not a part of the structure of a neuron, but rather the site of close proximity between two communicating neurons. There are two main types of synapses: electrical and chemical.
Electrical synapses
Electrical synapses are the simpler of the two types of synapses. In an electrical synapse, the main driver of communication between two neurons is a change in potential, and the carrier of charge is almost always an ion. The electrical synapse simply means that the cells share their cytoplasm with an adjacent cell. Electrical synapses are what Camillo Golgi imagined when he proposed the reticular theory of nervous system organization. Oftentimes an entire network of many hundreds of neurons is connected by these synapses.
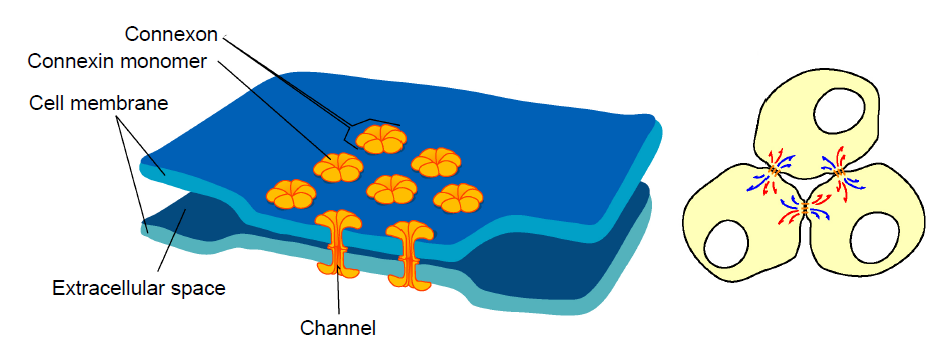
Figure 5.1 An electrical synapse exists between two closely-connected neurons. Cytoplasm passes between the two neurons through a protein complex called a connexon.
Imagine two neurons that are connected by an electrical synapse. First of all, both of them are complete cells on their own. Each one contains a complete plasma membrane surrounding the neuron, a nucleus, and all the individual organelles needed to carry out that cell’s basic life processes. Electrical synapses share the cytoplasm between the two connected cells, so ions, ATP, and larger signaling molecules and proteins are able to move between the two cells. For this to happen, there exists a specialized physical channel between the two that allows for the passage of cytoplasm called a connexon or hemichannel. Each hemichannel itself is made up of six transmembrane proteins called connexins (you can remember the difference between the channel and the individual protein because proteins often end with the letters -in).
When two connexons contact each other, they interact closely with each other and form the gap junction, which is the structure that connects neurons electrically.
Neurons that are connected by electrical synapses are remarkably close to each other. The synaptic gap between the two electrically connected neurons is about 3.5 nanometers. Logically, the neurons must be close since the hemichannels are like a physical “bridge” between the two cells.

Figure 5.2 Electrical synapses are similar to a skybridge that physically connects the cytoplasm between two neurons.
Electrical synapses are capable of passing information bidirectionally. This means that a signal does not always move sequentially from the presynaptic cell to the postsynaptic cell. Rather, ions and signaling molecules are free to move through the connexons in either direction. Also, each cell within an electrically- coupled network can receive inputs at any of the cells, making it able to detect several signals at once – the same way a huge satellite dish can detect more signals than a small dish.
Electrical synapses likely evolved because of evolutionary pressures that selected for speed. These synapses can pass signals as fast as electrical charges can move through an electrolyte-rich fluid like cytoplasm, which is almost instantaneous. Therefore, an escape reflex that is made up of communication across electrical synapses is advantageous for animals that need to escape predators. For example, crayfish exhibit a reflexive abdominal flexion response when exposed to threatening stimuli, causing the animal’s body to dart away from a threat within a fraction of a millisecond. Comparing across the phylogenetic tree, electrical synapses are often found in less complex organisms, including arthropods such as insects and crustaceans, where such reflexes are likely more critical for survival.
Another advantage of electrical synapses is that they can form a large network of interconnected neurons with synchronized activity. For example, neuroendocrine cells in the hypothalamus are connected by electrical synapses. When the “go” signal arrives, all the cells depolarize at once, which can result in the massive release of hormones into the bloodstream. A network can also cause sudden, powerful inhibition. Like an angry mob of people chanting, a network of electrical synapses connecting inhibitory interneurons allows the network to send an immediate “shut-down” signal under specific circumstances.
Clinical connection: Charcot-Marie-Tooth (CMT) disease
Charcot-Marie-Tooth (CMT) disease is a rare genetic disorder that damages parts of the peripheral nervous system including the motor nerves, resulting in muscle weakness and difficulty with walking, and the sensory nerves, causing some to experience abnormal sensations such as tingling or pain in their extremities. These symptoms are characteristic of signal transduction failure resulting from deficits in myelin. One type of connexin protein, called Cx32, is heavily expressed in Schwann cells, the glia that produce myelin in the PNS. Mutations in the gene that codes for Cx32 are associated with the X-linked form of CMT disease, and knocking- out the gene in experimental mice cause the mice to express similar symptoms as human CMT.
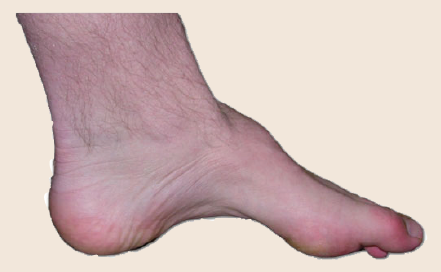
Figure 5.3 People with CMT disease often have abnormally shaped feet.
Chemical synapses
At a chemical synapse, a signaling molecule is released by the presynaptic cell to influence the postsynaptic cell. These signaling molecules, generally called neurotransmitters, are synthesized or stored by neurons. After being released, these neurotransmitters diffuse randomly across the synapse, where they are able to affect nearby neurons once the chemical binds to its corresponding receptor.
Figure 5.4 Chemical synapses are the site of close interaction between two neurons (left) or a motor neuron and a muscle fiber, which is called the neuromuscular junction (right).
Since chemical synapses do not rely on a direct physical protein “tunnel” to connect the two neurons, the distance between the two cells can be much larger. On average, a chemical synapse is a distance of about 20-40 nanometers, roughly a thousand times smaller than the diameter of a human hair.
A chemical synapse can pass a variety of signals, depending on the neurotransmitter and the receptor. For example, some signals are directly excitatory and allow positively charged cations to enter the neuron causing depolarization. Other signals are hyperpolarizing, and therefore inhibitory. And yet other signals are much more complex, inducing changes in protein expression that can modify cellular excitability over the course of minutes or hours.
Because of the complexity of the signals that chemical synapses can convey, evolutionary development through time has allowed for a tremendous variety of responses. Chemical synapses allow for fine-tuning of neural networks, giving these nervous systems a larger range of possibilities. The nervous systems of “higher” organisms like humans tend to have several chemical synapses since these signals are likely necessary for complex behaviors and cognition.
Many chemical synapses exist between the axon of one neuron and the dendrite of another neuron. One specific type of chemical synapse refers to the space between a motor neuron and muscle tissue, and this is called the neuromuscular junction, or NMJ. When the chemical signaling molecule acetylcholine (ACh) is released by the presynaptic motor neuron, it is detected by receptors that are expressed on the muscle. The release of ACh causes contraction of the muscle.
5.2 Properties of vesicles
Types of vesicles
Molecules of neurotransmitters are often stored in synaptic vesicles before being released. Synaptic vesicles are tiny spheres of lipids just like the cell membrane. These vesicles can be roughly characterized into one of two classes:
- Small vesicles. These vesicles have a diameter of 40 nanometers and a volume of about 30 cubic microns. Given the size of neurotransmitters, we can estimate at somewhere on the order of thousands to tens of thousands of molecules of neurotransmitter can be stored in each vesicle. Small vesicles store most of the neurotransmitters we most often think of, including glutamate, GABA, dopamine, and norepinephrine, for example. Small vesicles are almost always exclusive found in the axon terminals.
- Large dense-core vesicles. These vesicles are much larger than small vesicles, with a range of diameter from 100 to 250 nanometers. They store peptides such as dynorphin or enkephalin, which have chemical structures much larger than the other neurotransmitters. Since these peptides are packaged into their vesicles near the nucleus, large dense-core vesicles can be found in the cell bodies and all along the axons in addition to the axon terminal.
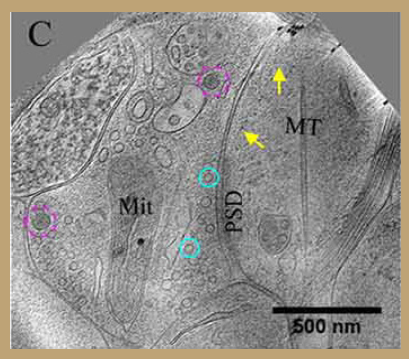
Fig 5.5 Electron microscope image showing small vesicles (cyan) and large dense core vesicles (magenta).
Loading of vesicles
Vesicles need to be filled with molecules of neurotransmitter before release into the synapse. In small vesicles, filling is only made possible through the action of giant transmembrane proteins called vesicular transporters. These are protein complexes that span the vesicular membranes, with one side facing the intracellular space and other facing the inside of the vesicle. Their main function is to take molecules of neurotransmitter from the intracellular space of the axon terminal and pump them into vesicles.
Many of the vesicular transporters are named based on the neurotransmitters that they are capable of recognizing and transporting. Some have a single substrate, such as vesicular GABA transporters (VGAT) which move GABA, vesicular glutamate transporters (VGluT) which move glutamate, and vesicular acetylcholine transporter (VAChT) which moves acetylcholine into vesicles. Others recognize a broad class of neurotransmitters, such as the vesicular monoamine transporters (VMATs), which are responsible for moving monoamines such as dopamine and serotonin into the vesicles.
Vesicular transporters are able to function because the interior of the vesicle is highly acidic compared to the interior of the cell. Vesicles have a high concentration of H+ ions (protons) because of the action of the transmembrane enzyme vesicular ATP-ase, or v-ATP-ase. These membrane-embedded proteins utilize the molecular energy contained in ATP to concentrate H+ ions in the intravesicular space. For each molecule of ATP used, one proton gets pumped into the vesicle.
Vesicular transporters pump molecules of neurotransmitter against their concentration gradients, which is an energetically difficult task. To have enough energy, the vesicular transporters use the high intravesicular concentration of H+ to move molecules of neurotransmitter across the vesicular membrane. When a proton moves from an area of high concentration to low concentration, energy is generated. The vesicular transporters use this energy to push neurotransmitter in. Because H+ ions move opposite of the neurotransmitter molecules, vesicular transporters are called antiporters. Transporters have slightly different stoichiometries, as it requires two protons to move a single molecule of dopamine, while the energy from a single proton is sufficient to transport GABA or glutamate.
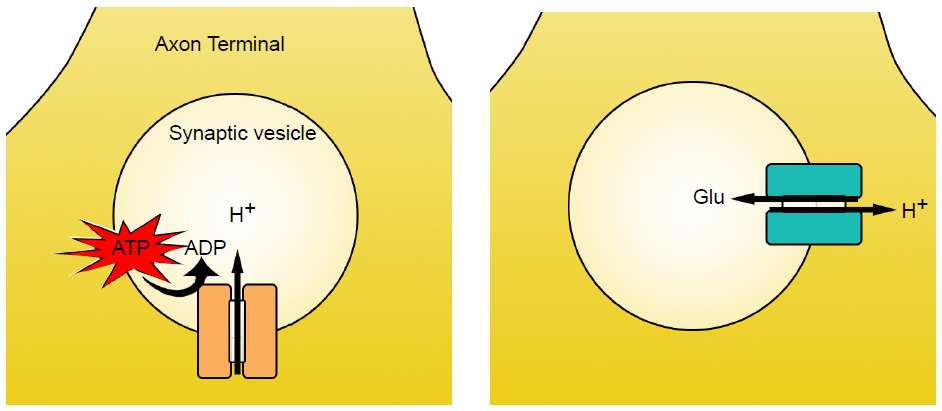 Fig 5.6 Synaptic vesicles in the axon terminal get filled by the action of two different vesicular transporter proteins. The v-ATP-ase uses energy to pump H+ into the vesicle against it’s concentration gradient (left). Then, a vesicular transporter such as vGluT use the movement of H+ down it’s concentration gradient to increase intravesicular concentration of neurotransmitter (right).
Fig 5.6 Synaptic vesicles in the axon terminal get filled by the action of two different vesicular transporter proteins. The v-ATP-ase uses energy to pump H+ into the vesicle against it’s concentration gradient (left). Then, a vesicular transporter such as vGluT use the movement of H+ down it’s concentration gradient to increase intravesicular concentration of neurotransmitter (right).
Location of vesicles
Synaptic vesicles can be found in one of three places at the axon terminal.
- Readily releasable pool (RRP). These vesicles are located close to the cell membrane at the axon terminal. In fact, many of them are already “docked,” meaning that their coat proteins are already interacting closely with the proteins on the inside of the cell membrane. When the depolarizing charge of an action potential reaches the terminal, these vesicles at the RRP are the first ones that fuse with the cell membrane and release their contents.
- Recycling pool. These vesicles are the ones that have been depleted due to release. They are currently in the process of being refilled or reloaded with neurotransmitter. They are farther from the cell membrane, and the protein machinery is not primed for release, so it requires a more intense stimulation to release the contents of these vesicles.
- Reserve pool. These vesicles are the farthest from the surface of the cell membrane, and most vesicles are held in this reserve pool. For these neurons to be released, very intense stimulation is required. Reserve pool vesicles may not even be recruited for release under physiological conditions.
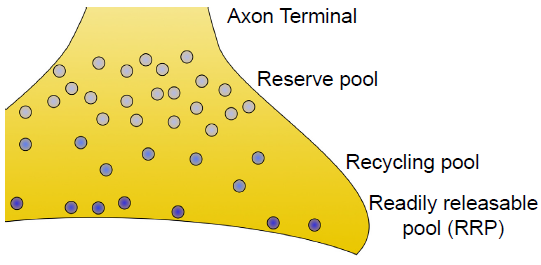
Fig 5.7 Axon terminals contain hundreds of vesicles roughly divided into three categories.
Release of vesicles
At a chemical synapse, the process of neurotransmitter release is very tightly regulated. If there were no mechanisms to control the release of chemicals at the synapse, nerve cells would deplete their entire stock of neurotransmitter. The signals that trigger muscle contraction at the NMJ would cause constant muscle tension. All sorts of brain signals would be active, and over-excitation would cause toxicity. Needless to say, regulated control of neurotransmitter release is a normal and essential part of nervous system function.
Regulation of release depends on several proteins that are important parts of the process. These proteins are often embedded within cell membranes of the vesicles or the neuronal membrane.
- V-SNAREs are the proteins expressed on vesicles (v for vesicle). Synaptobrevin and synaptotagmin are two specific v-SNARE proteins that are involved during synaptic release.
- T-SNAREs are proteins expressed on the cytoplasmic side of the axon terminal. The inside of the cytoplasm is the “target” for the vesicle (The t in t-SNARE). Syntaxin and synaptosomal nerve-associated protein 25, or SNAP-25 for short, are t-SNAREs that function during vesicular fusion.

Fig 5.8 v-SNARE proteins interact with t-SNARE proteins to allow for vesicular fusion and release of neurotransmitter.
Clinical connection: Botulism
Botulism is a deadly condition that results from exposure to the spores produced by the bacteria Clostridium botulinum. Toxic spores can be found in the soil, contaminated foods, or water. The toxin itself is one of the most potent agents known to man – exposure to concentrations as low as 2ng/kg is lethal. The most common symptoms include muscle weakness or paralysis, especially the muscles of the face or the limbs. For about 5% of people who develop botulism, death results from paralysis due to respiratory failure.
Botulinum toxin is known to selectively cleave the proteins that comprise the SNARE complex. There are a few specific types of botulinum toxin with slightly different intracellular targets, but the result is the same on the molecular level: prevention of vesicular fusion eliminating neurotransmitter signals.
Despite being one of the deadliest toxins so far identified, millions of people pay to have a preparation of toxin called “Botox” injected into their face. For most, the injection of Botox is a cosmetic procedure that can reduce the appearance of wrinkles by paralyzing the muscles. Botulinum toxin is also used medically for conditions resulting from excessive neurotransmitter release, such as muscle spasms, excessive sweating, or migraine.
Fusing of vesicles
The last step of neurotransmitter release is the fusing of the cell membrane. In order to release their chemical contents into the synapse, vesicles need to fuse with the cell membrane. As the vesicular membrane merges with the interior of the neuronal membrane, the contents of the vesicle become exposed to the extracellular space. Only then are the neurotransmitters capable of activating receptors.
One of the key proteins required for vesicular fusion is the vesicle-embedded V-SNARE synaptotagmin. This protein is capable of detecting elevated levels of Ca2+ in the axon terminal. As it turns out, an elevation of Ca2+ in the intracellular space is the “go ahead” signal that causes neurotransmitter release.
The concentration of intracellular calcium, generally in the range of 100 nM, is much lower than the concentration outside the cell. Embedded in the cell membrane of the axon terminals are transmembrane proteins called “voltage- gated calcium channels” or VGCCs. As their name suggests, they function very similarly to the voltage-gated sodium channels described in previous chapters: they are large protein complexes that normally remain closed, but when the surrounding neuronal membrane becomes depolarized, they physically change conformation and open up, allowing ions to move across the cell membrane. These VGCCs selectively pass only Ca2+ ions. The electrochemical gradient causes these Ca2+ ions to enter the cell.
As the change in membrane potential travels down the length of the axon (the action potential), it causes a depolarization at the terminal, triggering calcium entry via the VGCCs. Ca2+ at the terminal binds with synaptotagmin. The v-SNAREs and the t-SNAREs interact with one another in the presence of Ca2+, forming a molecular structure called a SNARE complex. The SNARE complex looks a lot like two twist ties that are wound tightly together. As they twist tighter together, it causes the vesicle membrane to approach the inside of the cell membrane, which results in vesicular fusion.
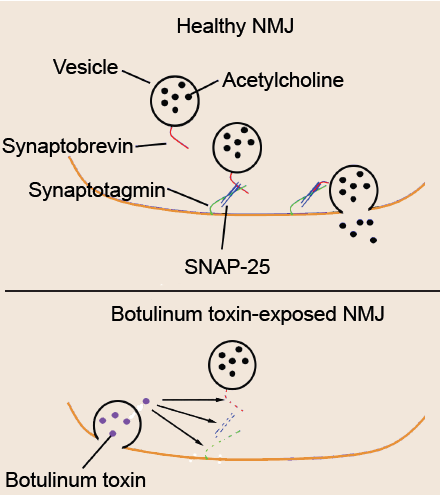
Fig 5.9 Botulinum toxin selectively cleaves vesicular fusion proteins, preventing acetylcholine from being released at the NMJ.
Vesicles are capable of fusing in at least two different ways.
- Full fusion. A vesicle that undergoes full fusion experiences total exocytosis. The vesicular membrane becomes completely integrated with the cellular membrane, and all of the neurotransmitter spills into the synapse.
- Kiss-and-run. This method of neurotransmitter release is incomplete fusion. The vesicle only partly connects with the interior surface of the cell membrane, and only a limited number of neurotransmitter molecules are able to enter the synapse via diffusion.
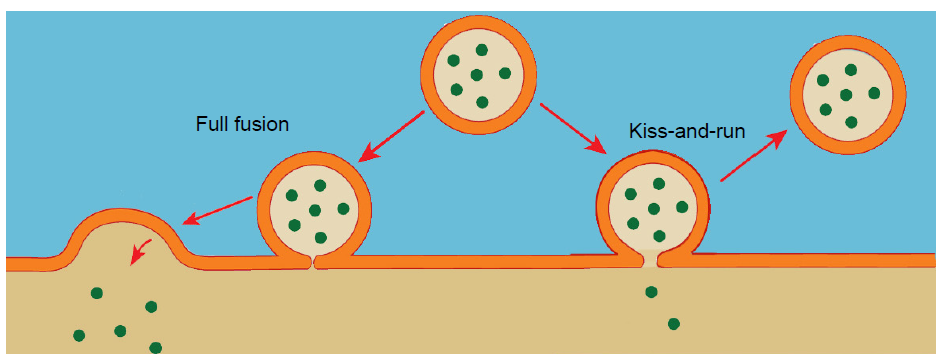
Fig 5.10 Synaptic vesicles either fuse completely (left) or partially in kiss-and-run (right).
5.3 Receptors
Receptors are proteins that are capable of sending a signal to change the function or activity of a neuron. Most receptors that function in neurotransmission are large transmembrane proteins. On the extracellular surface is a specific series of amino acid residues called the active site. The active site, also called the orthosteric site, is shaped to allow molecules of neurotransmitter to bind to the receptor.
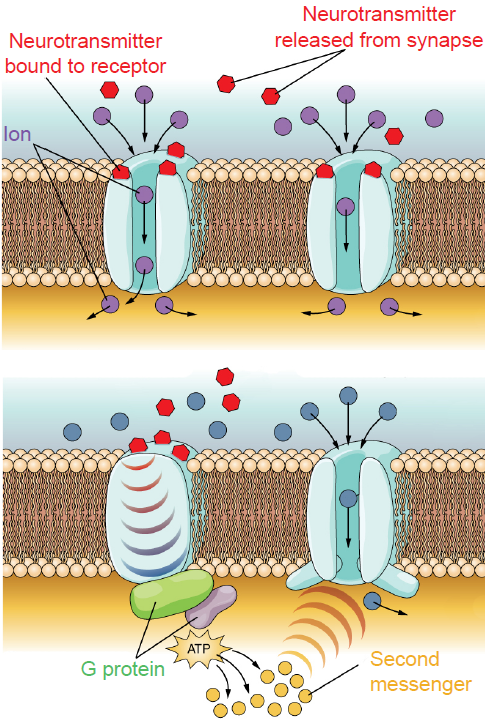
Fig 5.11 Ionotropic receptors (top) allow ion movement after receptor binding, while metabotropic receptors (bottom) trigger second messengers to induce signaling.
Receptors are classified into one of two main categories.
- Ionotropic receptors. Physically, ionotropic receptors are transmembrane proteins with a large-diameter pore through which ions can pass. These channels only open when a molecule of chemical binds to the active site on the extracellular side of the protein. These chemicals are also called ligands, and so ionotropic receptors are also called ligand-gated ion channels. Once a neurotransmitter activates the ionotropic receptor, ions will move based on the electrochemical gradient for that ion. As a result of ion movement, the cell’s membrane potential will change. For example, nicotinic acetylcholine receptors are ligand-gated sodium channels, so when these receptors are activated by a ligand like acetylcholine, sodium ions enter the cell causing depolarization and excitation.
Ionotropic receptors are able to induce a change in membrane potential very rapidly, on the scale of milliseconds.
Due to the nature of the amino acid residues that make up the pore of ionotropic receptors, they can be very selective for certain ions. For example, negatively charged residues lining the inside of the pore repel negatively charged Cl- ions while allowing positively charged cations to pass through the channel.
- Metabotropic receptors. These receptor complexes cause the cell to change its metabolism in a way that leads to either excitation or inhibition. Ions do not pass through these receptors. Instead, metabotropic receptors use the actions of G proteins, proteins which induce changes in neuronal excitability through the action of second messenger signaling molecules.
Physically, metabotropic receptors are transmembrane proteins that contain 7 alpha- helix motifs that pass through the cell membrane. The N-terminus of the protein is extracellular, and the protein “weaves” back and forth across the cell membrane, resulting in a protein with three extracellular loops and three intracellular loops. Because of this shape, these receptors are also called seven-transmembrane receptors, or 7-TM receptors.
Another name for these receptors is “G protein-coupled receptors”, or GPCRs. Metabotropic receptors are physically linked to proteins called G proteins, which exist on the inner surface of the cell membrane. Functionally speaking, these G proteins are capable of binding to molecules of guanosine triphosphate (GTP) or guanosine diphosphate (GDP). Chemically similar to ATP, GTP can function as a source of energy. G proteins themselves exhibit catalytic activity of GTP. This means that they are capable of breaking down GTP into the less-energetic GDP. When GTP is bound to the GPCR, the receptor is active. When this molecule is hydrolyzed into GDP, the receptor becomes inactive.
Some G proteins are heterotrimeric, meaning that they are made up of three different subunits, alpha, beta, and gamma. The GTP binding sites and catalytic sites are found on the alpha subunit, the largest of the three subunits.
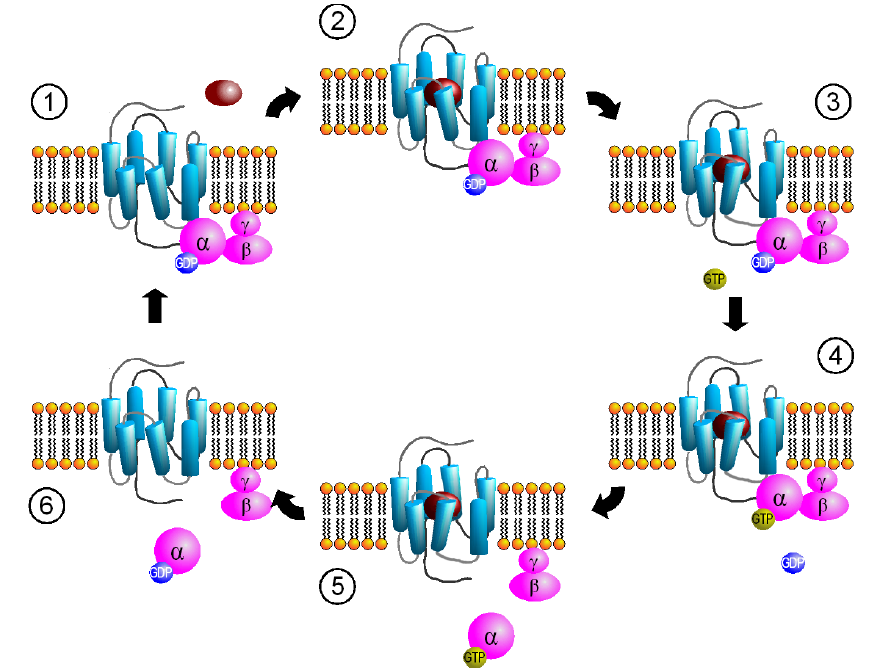
Fig 5.12 GPCRs signal via activation of the G protein attached to the intracellular side of the receptor.
Usually, the alpha subunit becomes soluble after activation, while the beta / gamma complex stays embedded in the neuronal membrane. The G alpha subunit exists in different varieties.
Gαs. When a neurotransmitter activates a GPCR coupled with the Gαs protein, the Gαs protein is excitatory (The “s” stands for stimulatory).
Binding of a ligand to the active site of Galphas-coupled GPCRs results in increased activity of the enzyme adenylate cyclase (AC). AC itself is an enzyme that creates a second messenger, a molecule called cyclic AMP (cAMP). Elevated levels of cAMP activate an enzyme called protein kinase A (PKA).
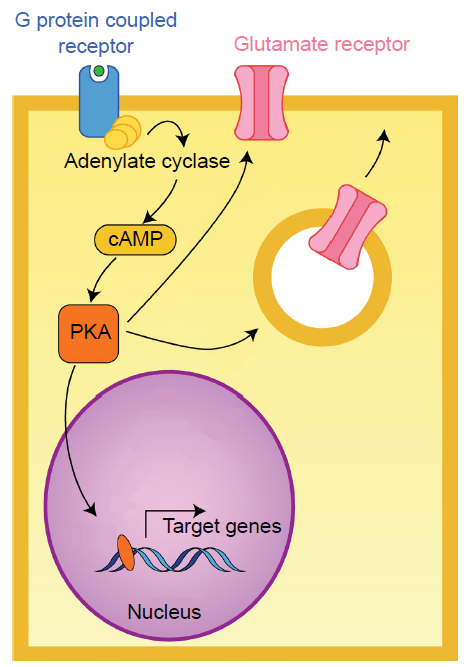
Fig 5.13 GPCRs that are coupled with Gαs are excitatory through adenylate cyclase signaling.
PKA is a kinase, an enzyme that is phosphorylates other proteins. The addition of a phosphate group onto a protein changes its properties dramatically. PKA phosphorylates protein targets that increase cell excitation. For example, one target of PKA activity is the intracellular side of certain glutamate receptors. Phosphorylation causes these receptors to stay open longer than normal when they are activated by a molecule of glutamate. This means that a single molecule of glutamate causes more excitation (passes more depolarizing current into the cell) in the presence of increased PKA activity. Targets of PKA activity also include the intracellular store of glutamate receptors. When phosphorylated, these receptors are trafficked to the neuronal membrane. An increase of receptors at the postsynaptic side leads to increased excitatory neurotransmission over a period of minutes and hours, representing one form of plasticity.
An even longer-term action of PKA is its ability to change the transcription of various genes, which can trigger the synthesis of proteins. Some genes downstream of PKA activity include the structural protein actin, important for morphological changes in neuronal structure, or ion channels which change neuronal responses to neurotransmitter release.
Gαi. A GPCR that is coupled with Gαi causes a decrease in excitability. In many ways, Gαi proteins serve the opposite function as Gαs proteins – the “i” stands for inhibitory.
Whereas activation of Gαs increases the action of AC, Gαi proteins decrease AC activity. Therefore, Gαi activation decreases the intracellular concentration of cAMP, in turn decreasing PKA activity. Given the function of PKA as a kinase that increases cellular excitation as described above, decreased PKA activity inhibits cellular activity through multiple mechanisms, some of which include decreased current through glutamate receptors, decreased trafficking of glutamate receptors to the presynaptic neuronal membrane, and decreased transcription of certain genes.
Gαq. Generally, Gαq is an excitatory G protein. Gαq uses a different signaling pathway compared to the PKA pathway that is downstream of Gαs or Gαi. Gαq protein activation leads to activity of the enzyme phospholipase C (PLC).
On a biomolecular level, PLC acts on the phospholipid membrane molecule phosphatidylinositol 4,5-bisphosphate (PIP2). PLC is a hydrolytic enzyme, and it breaks PIP2 into two separate second messenger molecules: the soluble inositol triphosphate (IP3) and the membrane-embedded diacylglycerol (DAG). One of the functions of IP3 is to liberate Ca2+ from intracellular stores, which elevates intracellular Ca2+ levels, depolarizing the cell and activating calcium-dependent processes, which are often excitatory. DAG activates protein kinase C (PKC), an enzyme with substrates that increase neurotransmitter release probability or decrease potassium channel conductance.
While the alpha subunits carry out a large part of GPCR-mediated changes in cellular excitation, the beta and gamma subunits also affect excitability. The beta and gamma subunits are bound together as a dimer, but they separate from the alpha subunit once the GPCR becomes activated. The beta-gamma complex can also function as a signaling molecule.
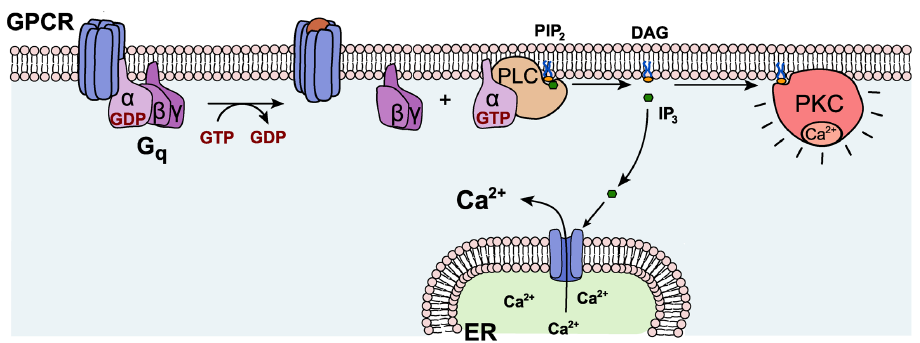
Fig 5.14 Gαq signals using PLC, which then produces two signaling molecules, IP3 and DAG.
Compared to ionotropic receptors, metabotropic receptors affect neuron activity on a slower time scale, on the range of milliseconds to seconds, and possibly even longer depending on the downstream mechanisms that are activated.
Presynaptic receptors
In the discussion of receptors, it is common to think of receptors as being expressed at the dendrites, embedded within the postsynaptic membrane. However, not all receptors are located here. Some receptors are presynaptic, meaning they can be found at the axon terminal. Presynaptic receptors are often inhibitory and serve a self-regulatory function. Presynaptically- expressed receptors that respond to the same neurotransmitter that is released are called autoreceptors.
5.4 Neurotransmitters
As described previously, neurotransmitters are the substances that are released at chemical synapses, and they are the signaling molecules that allow neurons to communicate with one another. To date, scientists have identified more than 100 neurotransmitters. Here, we will describe six classical neurotransmitters, their receptors, and their actions. Additionally, three atypical neurotransmitters will be introduced.
One important note to keep in mind as you think about neurotransmitters: the effect that a neurotransmitter has on a cell depends on the receptor. In other words, a neurotransmitter molecule can either excite or inhibit a neuron depending on the composition of receptors that are present. For example, glutamate is excitatory at most synapses in the nervous system. Glutamate exerts excitation by activating ionotropic glutamate receptors, which are ligand-gated cation channels. However, at one particular synapse in the eye, glutamate activates a metabotropic glutamate receptor that causes cellular inhibition.
Glutamate
Glutamate (Glu) is the main excitatory neurotransmitter used by the nervous system. Glutamate is the same as the amino acid glutamic acid. There is more glutamate per volume of brain tissue than any other neurotransmitter. Glutamatergic neurons are identified by the presence of the vesicular glutamate transporter (vGluT).
Glutamate can activate both ionotropic and metabotropic receptors. Ionotropic glutamate receptors are all ligand-gated cation channels, which makes them excitatory since they allow Na+ to move into the cell. Ionotropic glutamate receptors are generally subdivided into three classes, named after exogenous chemicals that can activate the receptor. AMPA receptors are Na+ channels, but some also allow Ca2+ entry. NMDA receptors allow both Na+ and Ca2+ to pass across the membrane. When the cell is at rest, NMDA receptors also have a large magnesium ion in the pore that blocks ion movement through the channel. The third category of ionotropic glutamate receptors is called kainate receptors, which are similar to AMPA receptors.
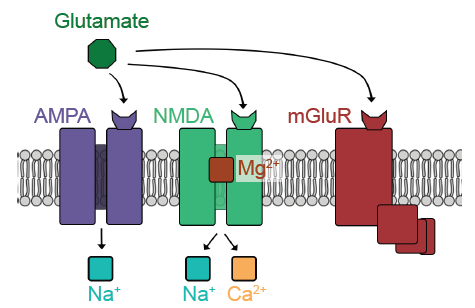
Fig 5.15 Glutamate is the main excitatory neurotransmitter in the nervous system, acting at different categories of receptors, three of which are shown below.
The metabotropic glutamate receptors (mGluRs) signal using different G proteins. There are a total of 8 of these mGluRs, classified into three groups, called Group I, Group II, and Group III. Group I are excitatory GPCRs which signal via Gq, while Group II and Group III are inhibitory via the Gi signal transduction pathway.
One theory proposes that excess signaling by glutamate can lead to neuronal death, a phenomenon called excitotoxicity. Of the various glutamate receptors, the NMDA receptor is most strongly implicated in contributing to excitotoxicity, since uncontrolled elevated levels of calcium can be deadly for neurons. Excitotoxicity is observed in a variety of disease states ranging from neurodegenerative disorders such as Parkinson’s disease, Alzheimer’s disease, and multiple sclerosis, but also in injury such as concussion or stroke.
GABA + glycine
Gamma-aminobutyric acid (GABA) is the main inhibitory neurotransmitter in the brain. According to one estimate, about 25% of neurons in the brain are GABA-ergic. Chemically speaking, GABA is remarkably similar to glutamate. In fact, GABA is synthesized from glutamate in a single step by the enzyme glutamic acid decarboxylase (GAD). GAD is often used as a biochemical marker for the presence of GABA-ergic neurons. Many interneurons use GABA as their chemical signaling molecule.
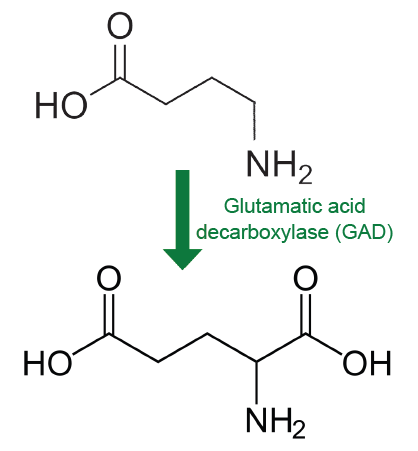
Fig 5.16 The inhibitory neurotransmitter GABA is synthesized from glutamate by the action of GAD.
The main action of GABA as an inhibitory neurotransmitter is to activate one of three main classes of receptors, called A, B, and C. GABAA receptors are ligand-gated chloride channels, so activating these ion channels causes Cl- flux, which opposes the ability for the cell to reach action potential threshold. GABAB and GABAC receptors are both metabotropic receptors that inhibit neuronal activity through the action of the Gi protein.
A neurotransmitter that is similar to GABA is glycine. Another small amino acid, glycine is mostly used by neurons of the spinal cord and brain stem. Glycine is also inhibitory , and acts at glycine receptors, which are ligand-gated chloride channels.
Dopamine
Dopamine (DA) is a biogenic amine derived from the amino acid tyrosine through the action of several enzymes. One in particular, tyrosine hydroxylase (TH), is the main marker that is used for identifying dopamine-producing neurons. Unlike glutamate or GABA, dopamine- producing neurons are not widely abundant in the brain. Instead, there are generally only a few patches of neurons that produce dopamine, most of which are found in the midbrain. Two areas include the ventral tegmental area and the substantia nigra.
There are five classes of dopamine receptors, named D1 through D5. All of them are metabotropic receptors. D1 and D5 are generally excitatory receptors, while D2, D3, and D4 are inhibitory receptors.
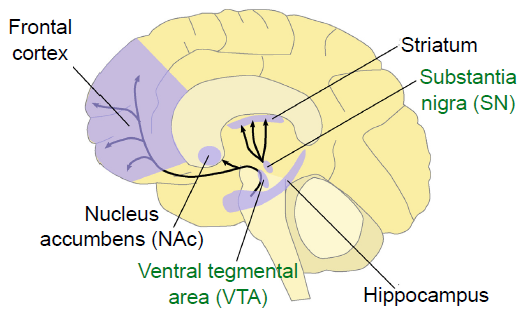
Fig 5.17 Brain dopamine is synthesized in two major midbrain nuclei, the VTA and SN, labeled in green.
Since Roy Wise’s theory proposed in the 1960s, DA has been known in pop culture as the “pleasure neurotransmitter” because of its involvement in the processing of reward and motivation. For example, if we use microdialysis (a technique to measure the concentration of chemicals) in the nucleus accumbens, dopamine levels spike in response to all sorts of pleasurable or rewarding stimuli: food, water, sex, sugar, and exposure to drugs of abuse. However, we now know that dopamine is much more complex than once believed. One theory suggests that dopamine elevation serves as a “learning signal” that causes us to pay attention to salient stimuli in the environment.
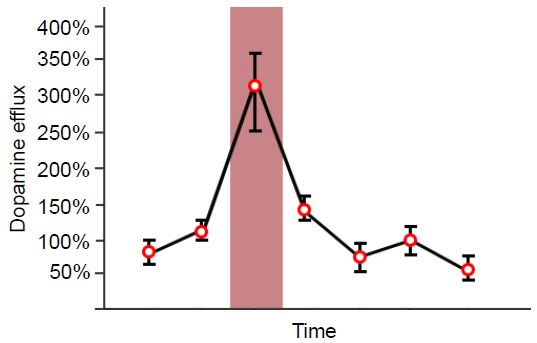
Fig 5.18 Levels of dopamine in the NAc rise when animals lever press for delicious foods, denoted in red.
DA is also needed for normal motor control. When dopamine-producing neurons in the substantia nigra pars compacta (SNpc) degenerate, as in Parkinson’s disease, a person develops the trademark symptoms: difficulties with motor control, resulting in a resting tremor, postural instability, and bradykinesia (slowness of movement). Reversing dopamine deficiency by introducing an exogenous source of dopamine is our current gold standard of treatment for PD.
Serotonin
Serotonin (5-HT) is a neurotransmitter that is derived from the dietary amino acid tryptophan. The enzyme tryptophan hydroxylase is the first step of serotonin biosynthesis and is often used as a marker to identify serotonergic neurons. As with dopamine, there are only a few areas of the brain that synthesize serotonin, the major one being the Raphe nucleus in the brain stem.
Receptors for serotonin have a wide variety of actions. We have identified seven major families of 5-HT receptors, which are designated by the number and subclasses which are designated by a letter. For example, the 5-HT2A receptor is metabotropic and excitatory via Gq signaling, while the 5-HT5 receptor is inhibitory via Gi signaling. Most of them are metabotropic receptors; only the 5-HT3 receptor is ionotropic.
Serotonin is heavily implicated in the regulation of mood and complex behavioral conditions. One of our most effective strategies for treating depression is the administration of a drug such as fluoxetine, which acts as a selective- serotonin reuptake inhibitor (SSRI). Pharmacologically, fluoxetine increases synaptic levels of serotonin by preventing reuptake, and for some people, this has a moderate ability to reverse depression. Serotonin signaling is also a target for drugs that treat anxiety, post- traumatic stress disorder, obsessive-compulsive disorder, schizophrenia, and more.
Clinical correlation: Parkinson’s disease (PD) and L-DOPA-induced dyskinesia (LID)
Parkinson’s disease is a debilitating neurodegenerative disorder that affects as many as 1% of all people aged 60 or older. Generally, PD is lethal within 16 years. By the time a patient presents to the clinic with motor dysfunction, they have already lost almost 60-80% of dopamine-producing neurons in this area!
For decades, clinicians have been using the biochemical precursor to dopamine, L-DOPA, to treat the symptoms. However, after chronic exposure to L-DOPA, the drug becomes less effective and has a shorter duration of therapeutic action. Worse still, frequent treatment can lead patients to develop hyperkinesias, an abnormal excess of movements. This iatrogenic disorder is called L-DOPA induced dyskinesia (LID).
Biomedical engineers have developed a promising non- drug approach to treating PD called deep brain stimulation. A small stimulating device is surgically implanted into the subthalamic nucleus of the brain. When this brain area is stimulated, neural circuits are recruited which restores normal motor control.

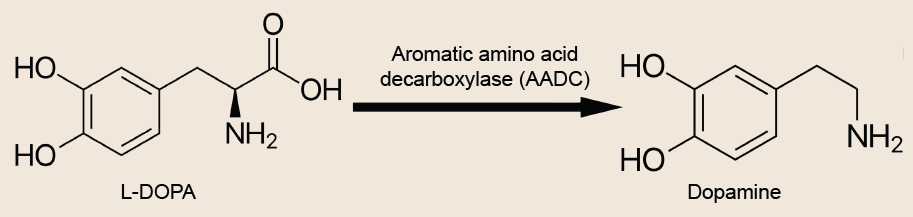
Fig 5.19 Patients with PD have characteristic changes in gait as a result of low dopamine (top). The current best pharmacological therapy is levodopa administration, which is the biochemical precursor to dopamine (bottom).
Acetylcholine
Acetylcholine (ACh) is a small molecule that is made by the enzyme choline acetyltransferase (ChAT), which chemically bonds a molecule of acetyl-CoA with a molecule of choline. The presence of ChAT in a neuron is used as a biochemical marker for neurons that produce acetylcholine.
ACh was the first neurotransmitter discovered and chemically isolated, a feat which earned two researchers the shared Nobel Prize in Physiology or Medicine in 1936. One of the two scientists, a German pharmacologist named Otto Loewi, stimulated the vagus nerve connected to an isolated frog heart, which caused the heart rate to slow down. When he put the surrounding solution on top of another heart, he observed that the second heart also slowed down, despite having no physical connection to the first heart. From this, he concluded that a chemical released by the vagus nerve is able to decrease heart rate. This chemical was first called Vagusstoff, the German word meaning Vagus substance. Today, we know it as acetylcholine.
ACh is able to act at ionotropic and metabotropic receptors, and activity at both receptor classes is essential for normal function. The ionotropic receptors of the nervous system are called nicotinic acetylcholine receptors (nAChRs) because they can be activated by nicotine in addition to acetylcholine. These ionotropic receptors are ligand-gated sodium channels and are therefore excitatory. On the other hand, the metabotropic receptors are called muscarinic acetylcholine receptors (mAChRs) since they are activated by the chemical muscarine found in some species of mushrooms. MAChRs can be coupled with either Gs or Gi, so they can be either excitatory or inhibitory.
ACh is the main neurotransmitter that the nervous system uses in order to communicate with the muscles at the neuromuscular junction (NMJ). Here, ACh is released by motor neurons, where it activates nicotinic acetylcholine receptors on muscle cells, causing them to constrict, or flex. On the other hand, muscarinic acetylcholine receptors are located in the heart, and their activation causes a decrease in heart rate (as Otto Loewi demonstrated with the isolated frog heart preparation.)
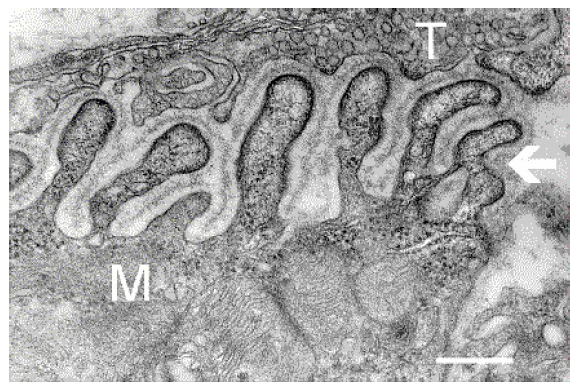
Fig 5.20 An electron microscope image of the neuromuscular junction showing vesicles (T, top) forming a synapse with the muscle cell (M, bottom). Acetylcholine is the main neurotransmitter used in muscle control at the PNS.
In the central nervous system, ACh is involved in a wide variety of processes, including attention and learning. One of the first theories to explain the symptoms of Alzheimer’s disease looked at a loss of ACh-producing neurons in the basal forebrain that become more severe as the disease worsens. It has since been demonstrated that Alzheimer’s disease is more complex than this early hypothesis.
Norepinephrine
Norepinephrine (NE) is a neurotransmitter that is synthesized from a molecule of dopamine by the enzyme dopamine beta-hydroxylase. Norepinephrine- producing cells are localized in the pons of the brain stem, a structure called the locus coeruleus. The locus coeruleus is very small, but these neurons send projections widely throughout the brain.
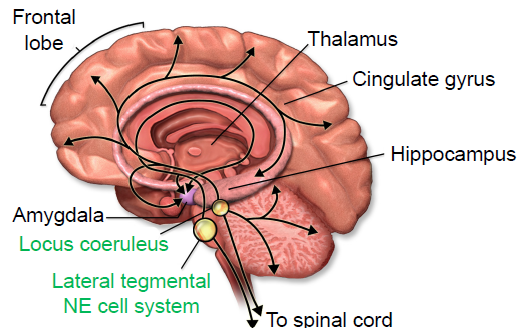
Fig 5.21 Norepinephrine in the brain is synthesized by small populations, but these cell bodies project widely across several other areas.
Outside of the brain, we think of norepinephrine as being responsible for triggering the sympathetic nervous system response of the body, the “fight-or-flight” reaction that prepares the body for physical activity in times of fear or acute stress. These norepinephrine-producing nerve cells reside in the sympathetic ganglia, a clump of nerve cells that run parallel to the spinal cord, one on each half of the body. These neurons project out towards the internal organs.
Receptors for NE are classified into two main categories, alpha or beta. There are subtypes within each category, giving us five major receptors for NE: alpha-1, alpha-2, beta- 1, beta-2, and beta-3. All five of these receptors are metabotropic, and some are excitatory while others are inhibitory. Our internal organs express these noradrenergic receptors. Clinically, the “beta blockers” are a class of drugs that inhibit beta-adrenergic receptors; the resulting action is a decrease in blood pressure. Conversely, some beta-agonists are used as bronchodilators for asthma.
Norepinephrine also functions in the brain to modulate behaviors including alertness and attention.
Atypical neurotransmitters
Although we generally think of neurotransmitters as neurochemicals that function as described above, there are a few atypical neurotransmitters that don’t quite fit the mold of the other chemical signals.
Neuropeptides
Neuropeptides are a class of large signaling molecules that some neurons synthesize. Neuropeptides are different from the traditional neurotransmitters because of their chemical size. Monoamines like DA, NE, or 5-HT have a molecular weight around 150- 200, while one of the smaller neuropeptides, enkephalin, has a molecular weight of 570. One of the largest, dynorphin, has a molecular weight greater than 2,000. Because of their large size, neuropeptides have to be packaged in dense- core vesicles very close to the site of production near the nucleus rather than in clear vesicles right at the terminal.
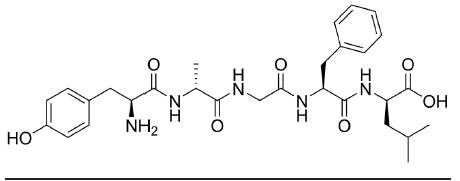
Fig 5.22 Enkephalin, one of the smaller neuropeptides, is very large compared to other neurotransmitters. Enkephalin is an agonist for both δ and μ opioid receptors.
Neuropeptides such as enkephalin and dynorphin are agonists at a class of receptors called the opioid receptors. These opioid receptors fall into four main types. The three classical opioid receptors are named using Greek letters: δ (delta), μ (mu), and κ (kappa), and the fourth class is the nociceptin receptor. All of these receptors are inhibitory metabotropic receptors which signal using the Gαi protein.
These receptors are expressed in several brain areas, but expression is particularly heavy in the periaqueductal gray, a midbrain area that functions to inhibit the sensation of pain. Drugs that activate the opioid receptors, like morphine, oxycontin, or fentanyl, are the most effective clinical treatments that we know of for acute pain. Unfortunately, these same drugs also represent a tremendous health risk, as opioid drugs can be lethal in overdose and have a high risk of substance use disorder.
Endocannabinoids (eCBs)
The eCBs are a class of lipid-based neurotransmitters. They are unusual neurochemicals in a few ways. Instead of sending information from the axon of one neuron to the dendrite of the next neuron (anterograde signaling), eCBs allow the postsynaptic dendritic component to communicate with the presynaptic axon terminal. Since they communicate information in the “opposite” direction of classic neurotransmitter signaling, eCBs are called retrograde signaling molecules. Secondly, eCBs are not packaged into vesicles and released by fusion processes. Instead, eCBs are synthesized de novo, meaning they get created right when they needed and used at that moment. The two most well-characterized eCBs in humans are called 2-AG and AEA.
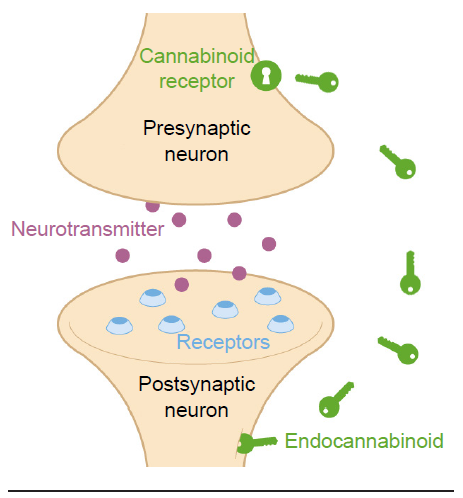
Fig 5.23 ECBs are synthesized from the postsynaptic cell membrane and signal through presynaptic cannabinoid receptors
ECBs activate one of two receptors, CB1 and CB2. Both of them are inhibitory metabotropic receptors that couple with Galphai. Generally, CB1 receptors are found in the nervous system, while the CB2 receptors are found elsewhere in the body, such as in the immune system.
The eCB system is widely used by various systems in the body. It is estimated that eCB receptors are the most abundant GPCRs in the whole body.
These substances were named because they are endogenous chemicals that are functionally similar to compounds found in plants of the genus Cannabis. One reason cannabis is used is because of its ability to interact with our eCB receptors.
Nitric oxide
The nervous system is capable of signaling via the gas nitric oxide (NO). This gasotransmitter is not stored in vesicles but rather is synthesized as it is needed. NO is formed when the amino acid arginine is degraded by the enzyme NO synthase (NOS).
Because NO is a gas, it easily permeates across cell membranes. Therefore, the receptors for NO do not need to be transmembrane proteins expressed on the cell surface. Instead, the receptor for NO is an intracellular receptor called soluble guanylate cyclase (sGC). SGC works through a signaling pathway that is different from other metabotropic receptors so far described. SGC is linked with the signaling molecule cyclic GMP (cGMP), which activates protein kinase G. PKG can either be excitatory or inhibitory, depending on the intracellular components.
Chapter summary
Neurons communicate with one another in a variety of ways. Anatomically, neurons are separated by a small extracellular gap called the synapse. This synapse may directly connect the intracellular cytoplasm, as in an electrical synapse. Alternatively, the gap may be much larger, and chemicals that get released and diffuse across the synapse in order to signal to the following neuron. These chemicals, the neurotransmitters, are stored in vesicles, tiny spheres that are located in the presynaptic axon terminal. The release of these chemicals is very closely regulated, and neurons have several mechanisms that regulate vesicular fusion.
Following release, the neurotransmitters diffuse across the synapse and can bind to the active site on transmembrane proteins called receptors. Upon binding a molecule of neurotransmitter, these receptors physically change shape, resulting in ion flow across the membrane (ionotropic receptors) or a change in the activity of intracellular signaling molecules (metabotropic receptors). Binding of neurotransmitter changes the excitability of neurons.
We have so far identified more than 100 neurotransmitters. Many of them are small molecules that are packaged in vesicles, which then diffuse from the presynaptic neuron to the postsynaptic neuron, such as acetylcholine, glutamate, or GABA. However, there are some atypical neurotransmitters such as neuropeptides, endocannabinoids, and nitric oxide that have different methods of communication.
The Open Neuroscience Initiative is funded by a grant from the Vincentian Endowment Fund of DePaul University.
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License.
Image Credits:
Cover: Image found at https://www.hippopx.com/en/people-letters-envelope-pen-dip-pen-writings-ink-62437
5.1 https://upload.wikimedia.org/wikipedia/commons/7/78/Gap_cell_junction_keys.svg modified by Austin Lim
5.2 https://upload.wikimedia.org/wikipedia/commons/e/e4/A_skybridge_at_IUPUI_campus.jpg
5.3 https://upload.wikimedia.org/wikipedia/commons/e/ed/Charcot-marie-tooth_foot.jpg
5.4 (Left) https://upload.wikimedia.org/wikipedia/commons/4/4c/Synapse_diag1.svg modified by Austin Lim (Right) https://commons.wikimedia.org/wiki/File:Synapse_diag4.png#file modified by Austin Lim
5.5 Tao C-L, Liu Y-T, Zhou ZH, Lau P-M and Bi G-Q (2018) Accumulation of Dense Core Vesicles in Hippocampal Synapses Following Chronic Inactivity. Front. Neuroanat. 12:48. doi: 10.3389/fnana.2018.00048
5.8 https://upload.wikimedia.org/wikipedia/commons/2/28/Opening_of_a_Fusion_Pore_during_Exocytosis.png modified by Austin Lim
5.9 https://upload.wikimedia.org/wikipedia/commons/1/1e/Botulinum_Toxin_Mechanism.png modified by Austin Lim
5.10 https://upload.wikimedia.org/wikipedia/commons/0/0c/0310_Exocytosis_cleaned.png modified by Austin Lim
5.11 https://upload.wikimedia.org/wikipedia/commons/e/e7/1226_Receptor_Types.jpg modified by Austin Lim
5.12 ttps://upload.wikimedia.org/wikipedia/commons/6/6d/GPCR-Zyklus.png
5.13 https://upload.wikimedia.org/wikipedia/commons/8/8f/CREB_cAMP_neuron_pathway.svg modified by Austin Lim
5.14 https://upload.wikimedia.org/wikipedia/commons/3/31/Activation_protein_kinase_C.svg modified by Austin Lim
5.17 https://upload.wikimedia.org/wikipedia/commons/d/de/Dopamine_pathways.svg modified by Austin Lim
5.18 Data from K.N. Segovia, M. Correa, J.D. Salamone, Slow phasic changes in nucleus accumbens dopamine release during fixed ratio acquisition: a microdialysis study, Neuroscience, Volume 196, 2011, Pages 178-188, ISSN 0306-4522, https://doi.org/10.1016/j.neuroscience.2011.07.078. Reprinted with permission 2/15/2020
5.19 https://upload.wikimedia.org/wikipedia/commons/4/44/Sir_William_Richard_Gowers_Parkinson_Disease_ sketch_1886.svg
5.20 https://upload.wikimedia.org/wikipedia/commons/a/aa/Electron_micrograph_of_neuromuscular_junction_%28cross- section%29.jpg
5.21 https://upload.wikimedia.org/wikipedia/commons/4/48/Norepinephrine_Part_1.png modified by Austin Lim
5.23 https://upload.wikimedia.org/wikipedia/commons/2/27/Sistema_endocannabinoide.png modified by Austin Lim
