3 Chapter 3: Cellular Anatomy of the Nervous System
COVER IMAGE
Austin Lim, PhD (DePaul University)
Editor: Katie Willis, PhD (University of Oklahoma)
For the majority of human history, the only way we were able to study the structure of the brain was with crude, butcher-like methods. Wait for a person to die, saw a giant hole in the top of the skull, take the brain out, and slice it into pieces to see if there was some correlation between the way the brain looks and the way they died. With these methods, only major changes in gross anatomy could be observed, such as that resulting from severe birth defects or trauma.
Brain analysis methods were enhanced by the scientific adoption of light microscopy. Naturalists in the mid-1600s such as Antonie van Leeuwenhoek, Jan Swammerdam, and Robert Hooke began looking at biological substances up close, and the brain proved to be a complex and interesting sample of tissue.
A major advancement in the study of neuronal morphology came about in the late 1800s. The Italian anatomist and biologist Camillo Golgi identified a shortcoming with the cellular analysis techniques of the time: structures in the CNS were impossible to distinguish from one another. The cells in the brain were so densely packed together, that it became difficult to identify which cellular material belonged to which cell. Golgi came up with a new technique using a silver compound that caused the silver to precipitate inside the cell membranes. However, not every cell took up the silver. Instead, only a small fraction of neurons, maybe 1% or even less, were completely stained in black, which stood out remarkably well against the light yellow background of the surrounding tissue. This reaction, initially called the “black reaction,” is now known as a “Golgi stain.” (Despite being more than a hundred years old, we currently don’t know the mechanism by which the silver stain is taken up into the neurons, or what determines why certain cells take the stain and others don’t.) Because of the great contrast between cell and background, every single part of the neuron was completely filled, allowing Golgi to do drawings of the morphology of this nervous tissue. Based on his staining results, Golgi supported the idea that the parts of the nervous system are all one very large, physically connected network. This idea was known as the reticular theory.
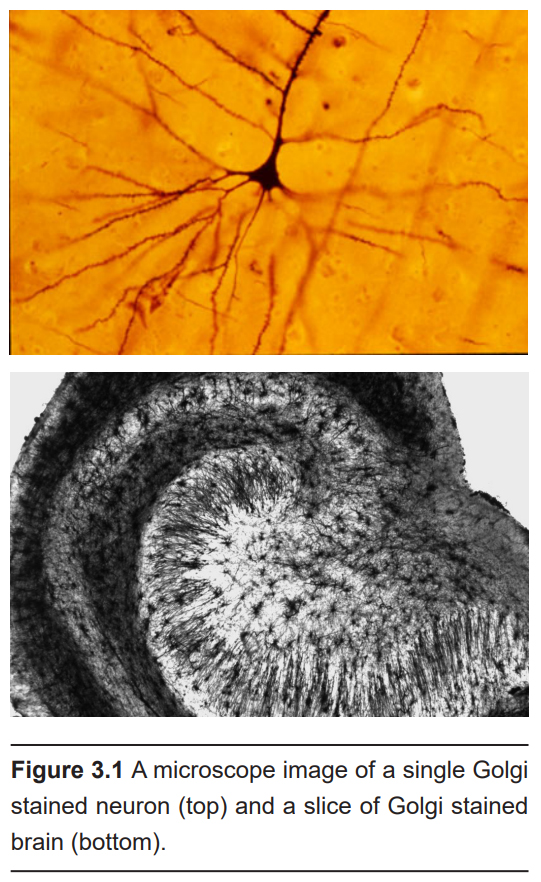
About 10 years later, the Spanish neuroanatomist Santiago Ramon y Cajal repeated some of Golgi’s staining experiments with other sections of nervous tissue. Looking at similar darkly-filled neurons, Cajal arrived at a different conclusion: the nervous system is not a giant net, but rather a series of individual units that are separated from one another physically. This idea came to be known as the neuron doctrine.
Both Golgi and Cajal were awarded the shared Nobel Prize in Physiology or Medicine in 1906 for their accomplishments in helping to understand “the structure of the nervous system.” Even though Cajal’s neuron doctrine was adopted widely by scientists, the elucidation of this organization would not have been made possible without Golgi’s development of the silver stain. The sharing of this prestigious award was ironic because of the many disagreements between the two scientists. Cajal commented on their relationship, saying:
“What a cruel irony of fate of pair, like Siamese twins united by the shoulders, scientific adversaries of such contrasting character!”

Cajal’s neuron doctrine was eventually given more support with the aid of modern techniques, like electron microscopy, that are capable of physically seeing the distance between two neurons. The neuron doctrine represents our current understanding of how the nervous system is organized, and this chapter is focused on describing the anatomical features of the nervous system at the cellular level.
Chapter 3 outline
3.1 Characteristics of neurons
3.2 Cellular anatomy of neurons
3.3 Cellular functions of glia
3.1 Characteristics of neurons
The main units of the nervous system are cells called neurons. Although neurons do have a variety of adaptations that make them unique from other types of cells in the body, they are still cells. Therefore, they contain all of the basic features of a typical mammalian cell. For example, they are made up of an aqueous cytoplasm bounded by a cell membrane. This cell membrane, also called a plasma membrane or lipid membrane, consists of a sheet of several individual molecules called phospholipids, which consist of two hydrophobic (water-fearing) tails and a hydrophilic (water-loving) end. These phospholipids arrange themselves into a bilayer with the hydrophilic tails touching each other and the hydrophilic sides facing the cytoplasm and the extracellular space, which are both mostly water. Because of the chemical properties of the cell membrane, it is very effective at keeping ions and charged molecules separated, while allowing small molecules like water and oxygen across the cell.
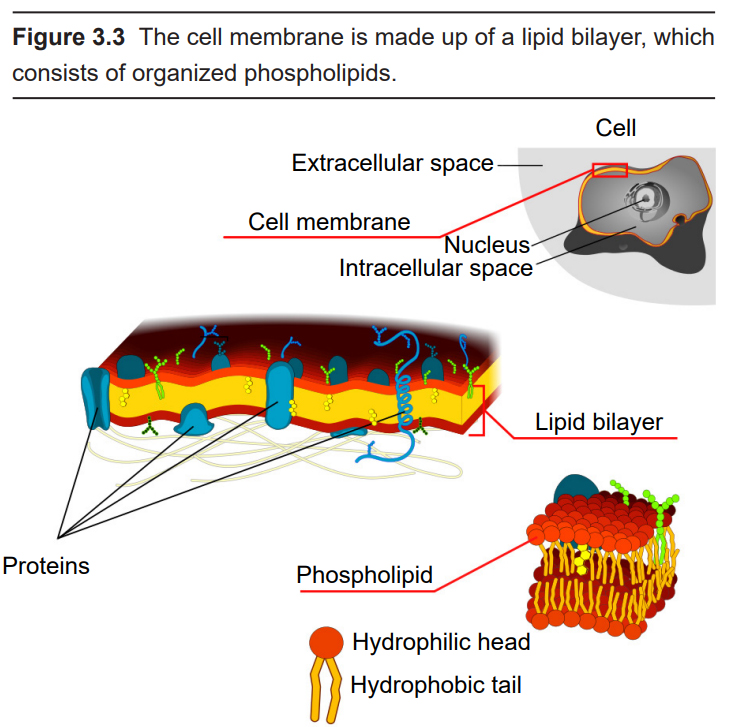
They also have all the organelles that you would see in other cell types, like a nucleus and mitochondria.
The number of neurons in the adult human brain, according to our current best estimate, is close to 86 billion. This number was calculated using a revolutionary technique, the isotropic fractionator or “brain soup” developed by Brazilian neuroanatomist Suzana Herculano-Houzel. To put this number in context, we have about 37 trillion cells in the whole body, so neurons in the brain make up about 0.2% of all cells in the body.
Below are some unique characteristics that neurons have in common.
Neurons are electroactive
The inside of most cells have a negative electrical charge compared to the solution outside of the cell. This difference between the electrical charges is called the membrane potential, or transmembrane potential. When a neuron is at rest, the amplitude of this charge may be somewhere between -50 millivolts (mV) and -90 mV. Often times, we say that in general the cell rests at around -70 mV. Membrane potential is abbreviated as Vm.
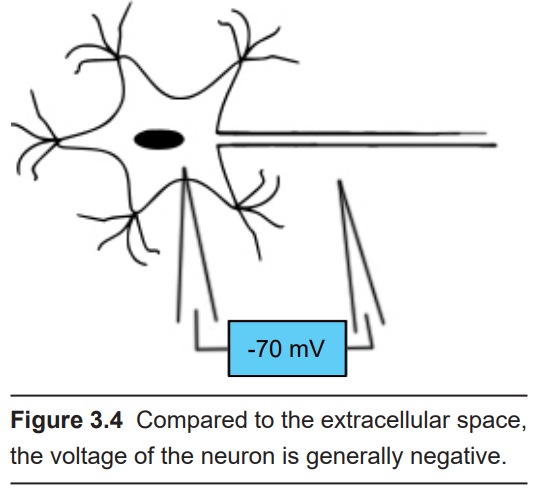
Most neurons do not spend their entire lives at -70 mV. Instead, neurons have special proteins embedded in their cell membranes that allow for charged ions, such as sodium or chloride, to move into or out of the cell. When a net positive charge enters into the cell, the Vm becomes more positive. Likewise, when net negative charge enters the cell, the membrane potential becomes more negative. The membrane potential of a neuron can go from -70 mV to +45 mV and back to -70 mV in as quickly as two milliseconds!
Neurons are not the only electroactive cells in the body. Our lives depend on cells in the heart that are also capable of changing in potential, as they respond to rhythmic electrical impulses that drive our heartbeat. This is why being struck by lightning may cause your heart to desynchronize in activity, and why a shock from a defibrillator can electrically “kick-start” the heart back to a meaningful pattern.
Neurons are specialized for rapid communication
Many cells are capable of sending and receiving chemical signals across long distances and time scales. But neurons are able to communicate with a combination of electrical and chemical signals in a matter of milliseconds. Additionally, the shape of neurons and the organization of the neurons on a microscopic level make them effective for sending signals in a very specific direction. Many neurons have an incoming receiving end and an outgoing sending end. The placement of one neuron next to the correct partner is very important, and many chemical signaling systems are in place to ensure that the developing nervous system is properly wired together.
Neurons are “forever” cells
We are constantly replacing non neuronal cells. For example, the cells in our bones replace themselves frequently at a rate of about 10% each year. Our body makes new skin cells to replace the dying skin cells on the surface so that we have a “new” skin every month. The cells along the inside of our stomachs, exposed to very harsh acidic conditions, get replaced about every week. About 100 million new red blood cells are created every minute!
On the other hand, the mature nervous system generally does not undergo much neurogenesis, the creation of new neurons. The neurons that we have after development are the ones that we will keep until we die, and this permanence of neuronal count makes them different from almost every other cell of the body. However, the idea of adult neurogenesis is a topic of debate among neuroscientists, since some areas like the olfactory system and the hippocampus display new nerve cell production.

…But, neurons can change
Even though new neurons are not created in most areas of the brain, neurons still have the capability to change in their structure and function. Some of these changes, such as physical changes to the structures of the input sites of the neurons, are believed to last for a lifetime. We use the word plasticity to describe the ability for the brain to alter its morphology (derived from Greek plastikos, meaning “capable of being shaped or molded” – think of plastic surgery, where a person changes their physical appearance; see chapter 13, Leaning and Memory).
Also, neurons do have the capacity to repair themselves to some extent. Neurons of the PNS may get injured or completely destroyed as a result of trauma to the body. Afterwards, those injured neurons can regrow to connect once again with their original partner. This regrowth seems to depend on a few chemical signals that the body produces, such as nerve growth factor and brain derived neurotrophic factor. However, this process is often very slow, and does not always successfully restore the nervous system to the way it was pre-injury.
3.2 Cellular anatomy of neurons
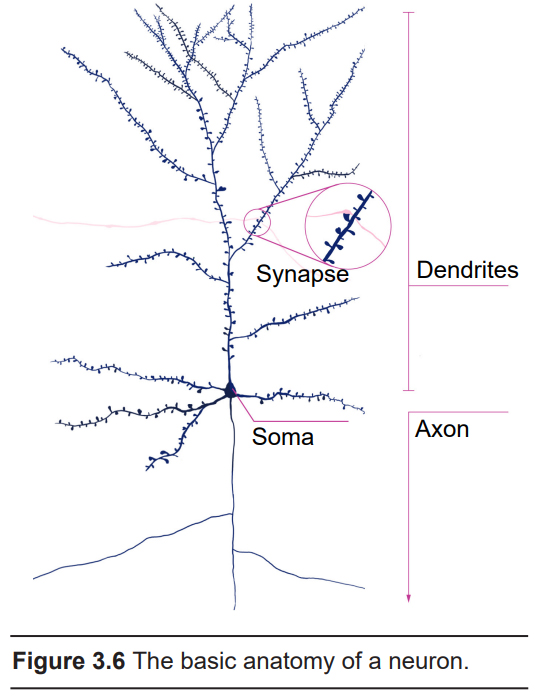
The main function of neurons is to use changes in electrical properties in order to communicate with connected cells. This communication usually moves in one direction, and we will use this pathway as an outline for discussing the anatomical structures of the neurons.
Dendrites
Information first enters the neuron through the dendrites, the branch-like extensions that protrude from the cell body. The word dendrite originates from the ancient Greek word “dendro-”, meaning “tree” (think of rhododendron). Dendrites look like the branches of a tree: they reach outward away from the center of the cell body, generally getting thinner the farther away you look.

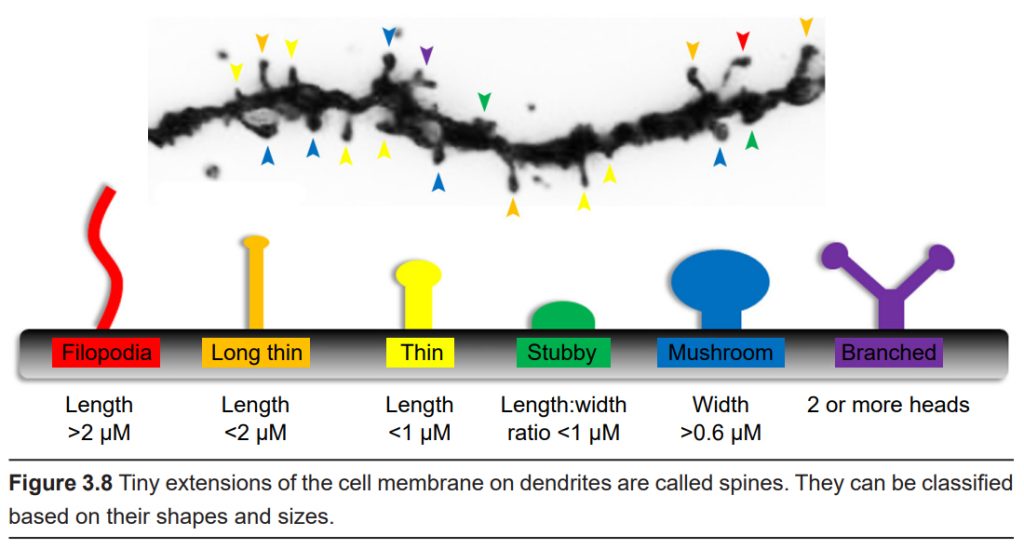
Along the dendrites of some neurons, it is possible to see tiny protrusions of cell membrane that stick out from the main dendrite. These bumps are called spines. Spines can roughly be classified based on their approximate shape; thin, mushroom, and stubby being some of the more easily recognizable forms. A spine may be about 100 nm in diameter, making it smaller than the wavelength of visible light, with a total volume of about 0.1 femtoliter – one ten-quadrillionth of a liter!
Chemical signals released by another cell are received by the dendritic spines, and so each spine may represent an input site of communication. Some cells, like a pyramidal neuron in the hippocampus, may have more than 30,000 spines, indicating that one cell may detect information from several incoming cells.
We believe that spines are one of the most important sites where the nervous system is able to change. For example, neurons change shape after exposure to various environmental conditions, such as stress or exposure to drugs. Tiny changes to the surface of the neuron at the level of dendritic spines is an example of plasticity. Dendritic plasticity is thought to underlie the reason that we can learn new facts or maintain memories about our childhood over long periods of time. Some set of tiny, submicroscopic changes to the morphology of dendritic spines may represent a single complex memory that you form.
A neuron does not need spines for receiving information or for plasticity to take place. Many cells lack spines, but are still capable of permanently changing. The input site may be anywhere along the dendrite, or even at the cell body, the “center” of the neuron.
Cell body (soma)
Information that arrives through the many dendrites of a neuron eventually filters into the cell body, or the soma, of the neuron. Somata (plural of soma) vary in size across different types of neurons, the largest somata belonging to the Betz cells of the motor cortex with an area upwards of 100 microns in diameter: about the size of a single grain of salt. On the opposite end of the spectrum, granule cells of the cerebellum are so densely packed so that they make up more 70% of all neurons in the brain, maybe 4 microns in diameter.
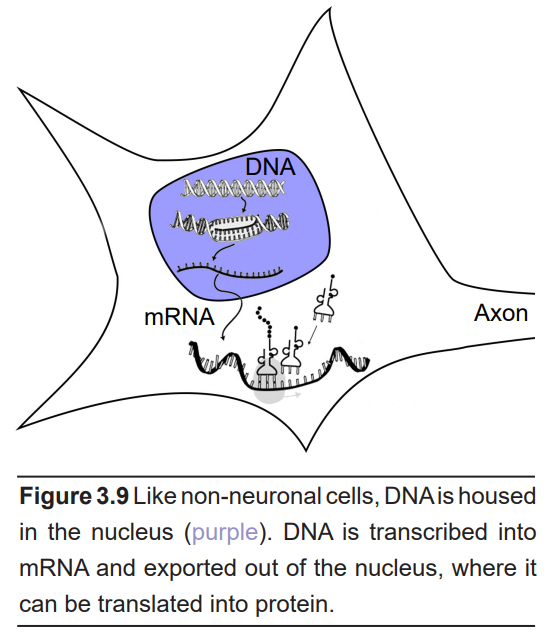
The cell body contains many of the organelles that are essential for the production of proteins that the neuron needs. Many of these organelles are not unique to neurons and can be found in other cell types. The most apparent organelle visible under high magnification (400x) is the nucleus, which houses DNA and other genetic material. From our understanding of the central dogma of molecular biology, this DNA is transcribed into a string of single-stranded genetic code called messenger RNA (mRNA), which is exported out of the nucleus. This mRNA is then used as a guide for the synthesis of proteins.
The next step of protein creation depends on organelles that are adjacent the nucleus. Physically continuous with the membrane that surrounds the nucleus is a folded membranous organelle called the endoplasmic reticulum (ER). Attached to the ER are several ribosomes, which are the molecular machines that read the mRNA and translate that code into proteins. The Golgi apparatus are layers of folded plasma membranes that function in transport. They are found near the nucleus, although small protrusions of these organelles may reach into the other parts of the neuron.
Axon
The axon is the main output extension of the neuron. While neurons only have a single axon extending from the cell body, this axon can branch several times after exiting the soma. In branching, an axon from a single neuron is able to communicate with many other neurons at the same time. Axons are usually thinner than dendrites, some being only a micron in diameter. Several axons can bundle and travel together; these are nerves. Axons can be very long; the longest axon in the human body is part of the sciatic nerve that runs from the posterior end of the spinal cord down the leg to control the muscles of the big toe.
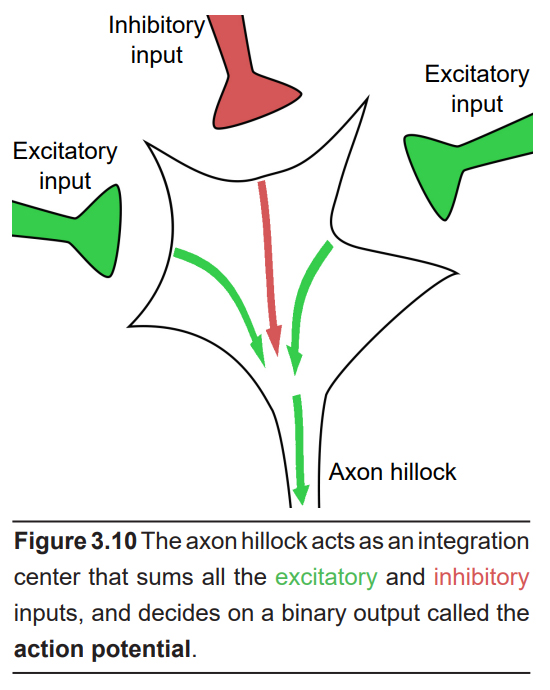
The first section of the axon, the junction between the cell body and the beginning of the projection, is a patch of axonal membrane with very unique characteristics. This section is called the axon hillock. With respect to signal transduction, the axon hillock works as the integrative center of the neuron. It is responsible for deciding whether to pass a signal onto the next cell. The cell membrane of the hillock performs a complex set of “cellular arithmetic” that weighs all of the incoming signals: excitatory, inhibitory, and modulatory signals. After all of the calculations have been performed, the membrane either sends a signal or not.
At the end of each branch of the axon is a small swelling, which is called the axon terminal or terminal bouton. The terminal is the part of the neuron that is specialized for the production and release of the neurotransmitters that are used for communication between neurons. One subarea of specific interest in the axon terminal is a small patch of membrane called the active zone. Embedded in the cell membrane at the active zone is a variety of proteins that are important for the process of neurotransmitter release.

Inside the axon
Most of the proteins synthesized in neurons are created in the cell body very close to the nucleus, where the mRNA that is exported from the nucleus is able to easily interact with the rough endoplasmic reticulum and the ribosomes. But, some of these proteins are needed far away from the cell soma, at the axon terminal, for example. Neurons need some system of transport system that can move newly created proteins to where they need to go. Inside the length of the axon runs an organelle called microtubules, which function like a molecular railway for proteins. The cells use motor-like proteins that can carry other proteins along the microtubules. When substances are transported away from the cell body, it is called anterograde transport, while the process of moving proteins towards the cell body is retrograde transport.

Another important organelle that is found along the length of the axon are neurofilaments. These organelles are made up of several different proteins that serve as a cellular “scaffolding” that helps keep the structure of the axon intact. Mature neurons can be very dense with neurofilaments, which increases the diameter of the axon.
Within the terminal are a number of vesicles, small spherical “packages” made of cell membrane that are coated in special proteins. Within these vesicles are the molecules that the neurons use for chemical communication. When the action potential travels down the axon and reaches the axon terminal, the cell membrane changes in electric charge, and this causes vesicles to fuse with the inner membrane of the neuron. During fusion, the vesicular membrane merges with the cell membrane at the axon terminal, causing the contents of the vesicle to be released outside the cell.
Outside the axon
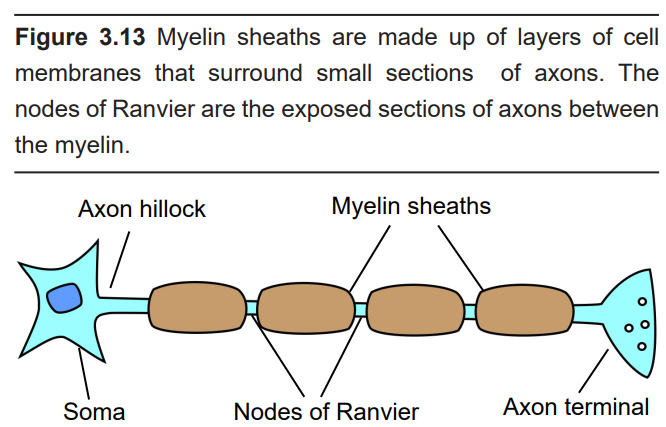
Some neurons have a special modification surrounding their axon called a myelin sheath. Myelin is comprised of several tightly-wrapped layers of cell membrane that encompasses a short section of the axon. Myelin might be wrapped as many as 250 or 300 times around a single section of axon. Myelin does not extend fully enclose the entire length of the axon from soma to terminal, but rather, surrounds short sections at a time. The spaces of exposed axon between each section of myelin are called nodes of Ranvier. On average, these nodes are about 1 micron long.
Myelin serves a few functional purposes. Myelin increases the speed by which an electrical signal is transmitted. Some of the most heavily myelinated axons are able to send signals up to 120 meters per second (almost 270 miles per hour, faster than a Formula 1 racer.) Myelin also increases the effective thickness of the cell membrane along the axon. In doing so, myelin acts as an insulator that causes signals to more reliably be passed down the axon.
Synapse
The synapse is the physical distance that separates two neurons. In agreement with Cajal’s neuron doctrine, the nervous system is not made up of a single cell with a giant, shared cytoplasm, but rather a series of neurons in close proximity, separated by a gap of extracellular space.
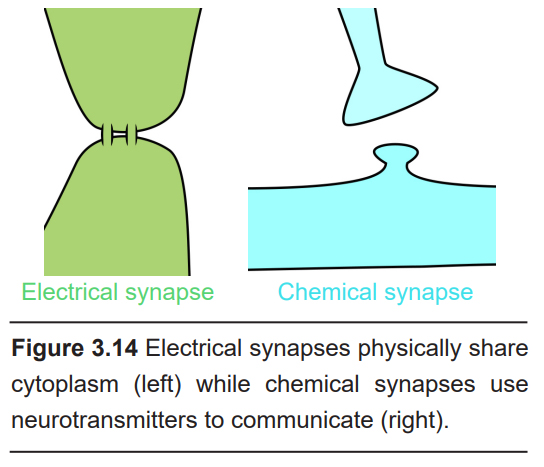
This distance between two cells can vary depending on the nature of the synapse.
An electrical synapse may be less than 5 nanometers apart. Cells connected by electrical synapses share cytoplasm but have two separate cell membranes. On the other hand, a chemical synapse is a larger distance, about 15 – 40 nm across. Adjacent neurons connected by chemical synapses do not share cytoplasm.
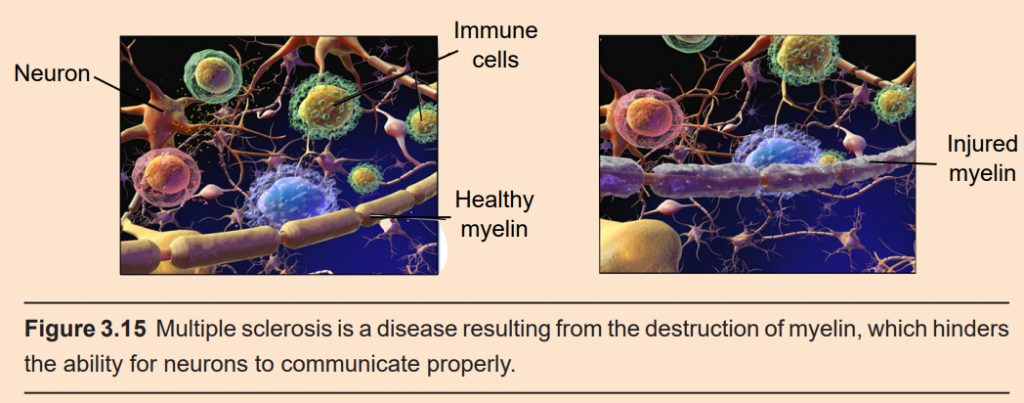
Clinical connection: Multiple sclerosis
Multiple sclerosis (MS) is a disease that results from destruction of myelin in the CNS. When myelin is damaged, signals do not reliably propagate from the brain to the body, from the body to the brain, or between areas of the brain. MS damages myelin at both descending and ascending neurons, so a person with MS might experience muscle weakness, poor balance, and muscle spasms (efferent motor neurons) as well as numbness and pain (afferent motor neurons). It can also affect neuronal signaling in the brain, causing symptoms such as cognitive impairment, vision loss, and changes in affect leading to depression.
Multiple sclerosis is a common neurological disorder that affects some 2.5 million people worldwide. MS typically presents in adulthood, usually between 20- 50 years old. Unfortunately, there is currently no known cure for MS. Therapy is focused on either slowing the progression of the disease, helping patients recover after an attack, or decreasing the severity of the symptoms.
While MS has the potential to be debilitating and painful, the disease itself is not necessarily lethal, and MS only decreases life expectancy by 5-10 years on average. People with MS are just as likely to die from natural causes as much as a neurotypical person.
One of the leading theories about the cause of MS is a faulty immune system. A normally functioning immune system identifies and destroys foreign pathogens. Sometimes, this system makes a mistake, and the immune response recognizes normal parts of the body as being a “foreign object”, causing the body to destroy itself. This is called an autoimmune disorder. In the case of MS, it is believed that the immune system identifies and targets myelin for destruction, leading to demyelination. Although immunosuppressants may slow the progression of MS, these drugs increase the risk of a person developing an illness that a healthy immune system would be able to prevent.
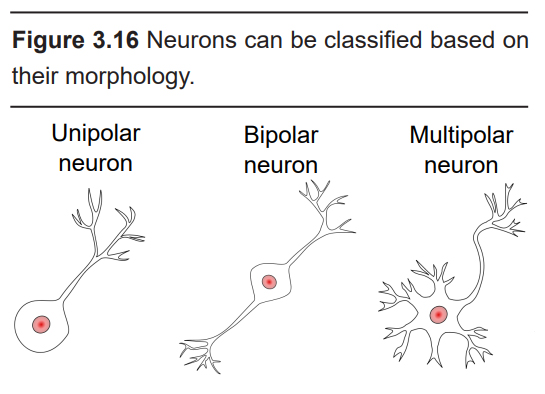
Classifications of neurons
Neurons have a wide variety of shapes and can be divided roughly into three different classes depending on their morphology. In order of increasing morphological complexity, they are:
- Unipolar cell
Unipolar cells have a single cellular extension coming off the soma that acts as both the receiving and the sending end. Unipolar cells are very common among invertebrates; humans do not have unipolar neurons.
- Bipolar cell
A bipolar cell, as can be implied from the name, has a single dendrite and a single axon. They are not very common, but can be found in human sensory systems, such as in the eye or in the signaling pathway connecting the ear to the brain.
- Multipolar cell
Multipolar neurons have several dendrites and one single axon. These cells are the most common among all the neurons in the human nervous system. Most drawings of neurons you will see in this book are multipolar cells.
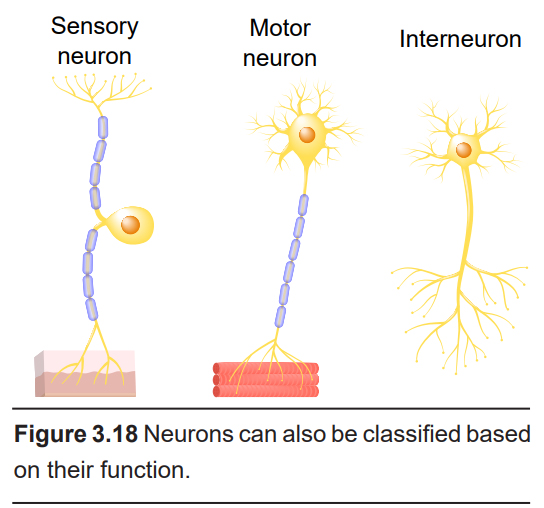
In addition to classifying neurons based on their morphology, neurons can also be divided based on their functions.
- Sensory
Sensory neurons are the afferent neurons that are responsible for obtaining information about either the outside world or the internal environment and passing that information towards the CNS. Sensory neurons can detect a variety of stimuli, ranging from photons of light (visual system) or chemicals floating in the air (olfactory system), to carbon dioxide levels in the blood (chemoreceptors) or stretching of the muscles (spindle receptors). Since they are highly specialized for detecting different types of stimuli, sensory neurons have a large variety of shapes and structures.
- Motor
Motor neurons, or motoneurons, carry signals from the CNS to the body. There are two main types of motor neurons. Somatic motor neurons control skeletal muscle movement, such as flexing or extending the muscles of the arm. They release neurotransmitter directly onto muscles. Autonomic motor neurons, on the other hand, release neurotransmitter onto a clump of neurons outside of the CNS called the autonomic ganglion, which then signal to the smooth muscle, cardiac muscles, or glands.
- Interneuron
Interneurons exist as a relay between the sensory or motor neurons and the CNS, or between each other. They represent a very broad class of neurons, and make up an important part of reflex circuits, such as the knee-jerk response.
The above classifications are only a rough guideline for separating neurons. As with most biologists, neuroscientists enjoy classifying cells based on their properties. It is almost always preferable to use the most specific name possible in identifying neurons. Many cells have been identified based on the way they look (chandelier cells), some named on the neurotransmitter released (cholinergic interneurons), some named by where they are found (cerebellar granule cell), and yet others named based on the person who discovered them (Purkinje cells).
Visualizing the synapse
The original microscopes that van Leeuwenhoek used to visualize cells and microorganisms relied on the transmission of light to see objects up close. For us to be able to see objects, light needs to bounce off the intended target. Light particles travel in a wave, and if the object we intend to visualize is smaller than the wave, then the wave would pass right over the object. The shortest wavelength of visible light is around 380 nanometers, so light microscopy is limited to visualizing objects in the micrometer or larger range – a 2,000x magnification with the best possible lenses.
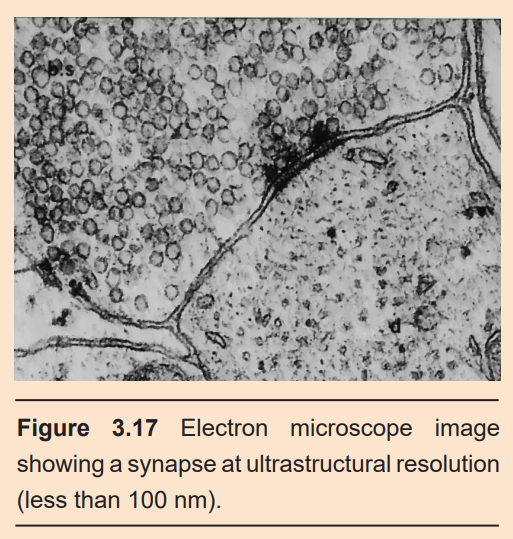
In the early 1950s, a group of scientists developed a technique called electron microscopy (EM). In EM, a beam of electrons is aimed at a sample in a vacuum, and the reflection of those electrons can be collected and detected with a computer. Since electrons have a much shorter wavelength than visible light, you are able to resolve objects as small as 50 picometers – more than a 10,000,000x magnification!
Using EM, you are able to get ultrastructural resolution of the synapse. In two adjacent neurons, EM allows you to distinguish the boundaries of both neurons, and to clearly see the synapse in between. You can also see the vesicles contained in the axon terminal, sometimes in the middle of fusion.
3.3 Cellular functions of glia
Although most of neuroscience is concerned with understanding the functions of neurons, there are other cells in the nervous system that are just as interesting. These cells are grouped together under the umbrella classification of glia. Historically, when these non-neuronal cells were visualized under the microscope, the histologists and anatomists had no idea about their function. They were seen all around the neurons, so the assumption was that these cells were structural elements, a sort of living glue that held the nervous together. Today, we know that these glia serve all variety of functions; unfortunately, the misnomer “glia,” derived from the Latin word for glue, is still used to describe these non-neuronal components of the nervous system. We estimate that the brain has roughly an equal number of glial cells and neurons—86 billion of each.
There are many different classes of glia, but we will focus on five types.
- Astrocytes
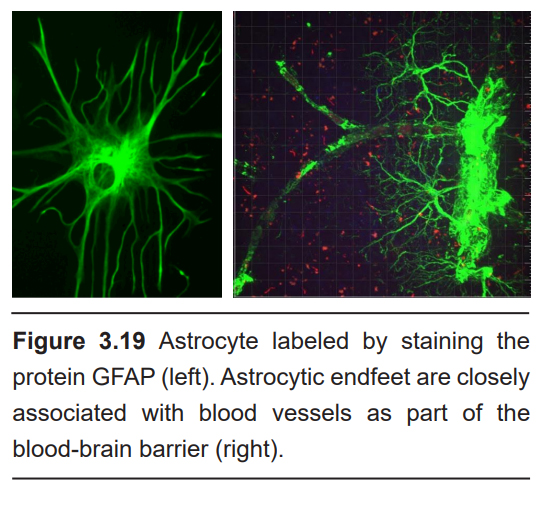
Astrocytes are named for their characteristic star-shaped morphology. Astrocytes have a dense expression of the protein glial fibrillary acidic protein (GFAP), and this protein is often used as a marker for differentiating astrocytes from other cell populations.
One of the main functions of astrocytes in the brain is to help maintain the blood-brain barrier. At the end of the extensions of the astrocyte are protrusions called “endfeet.” These endfeet are often wrapped around the endothelial cells that surround the blood vessels. The endfeet release important biological compounds that allow the endothelial cells to remain healthy as they function in maintaining the blood-brain barrier.
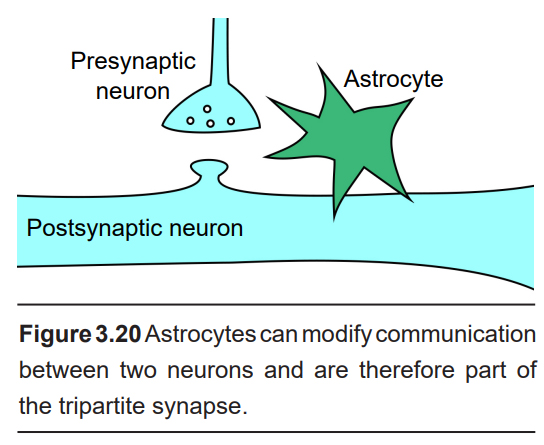
Astrocytes are also very closely associated with synapses. Astrocytes have a very dense expression of proteins on their cell surface that can transport molecules of the neurotransmitter glutamate, for example, inside the astrocyte. By acting as a glutamate “sponge,” astrocytes are able to decrease the strength of a glutamate signal. Through a similar uptake mechanism, astrocytes can also affect the extracellular concentration of ions such as potassium, which then has an influence on cellular excitability. Because of these interactions between astrocytes and neurotransmission at the synapse, we use the phrase tripartite synapse to refer to the three components of a synapse: The presynaptic neuron, the postsynaptic neuron, and the astrocyte.
Astrocytes also synthesize and produce a variety of trophic factors, which are helper molecular signals that serve several different functions. For one, trophic factors signal to neurons that the neuron should continue to live, or that specific synapses should be maintained. They help guide the neurons as they reach out, forming synapses where appropriate.
- Oligodendrocytes
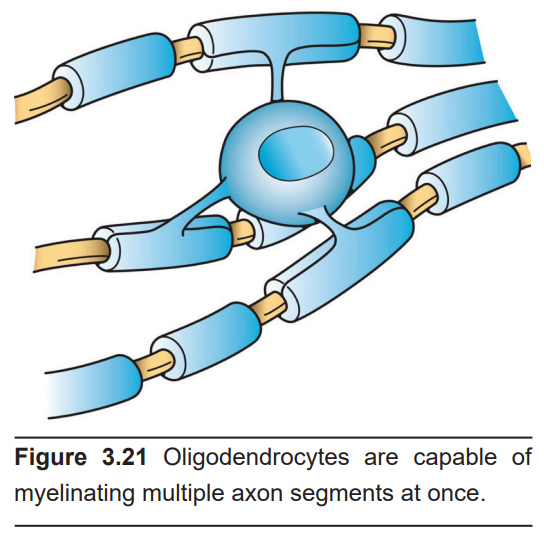
Oligodendrocytes only exist in the CNS. Their name is derived from their morphology. Oligo- refers to “a small number” (think oligarchy, a government ruled by a few people), and dendro- refers to “tree” (like a rhododendron). Each oligodendrocyte has a few branches that reach away from the cell body.
The main function of the oligodendrocytes is to add a layer of myelin around the axons of nearby CNS neurons. A single oligodendrocyte is able to myelinate up to 50 segments of axons. As cells that produce myelin, they are responsible for increasing the conduction speed of nearby neurons as they send signals. When the oligodendrocytes begin myelinating, they are able to produce almost 3 times their weight in membrane per day. By maturity, the oligodendrocyte supports cell membrane that is 100 times the weight of the cell. Due to the need to support such large amounts of myelin, it’s estimated that they have the highest metabolic rate of any type of cell in the brain.
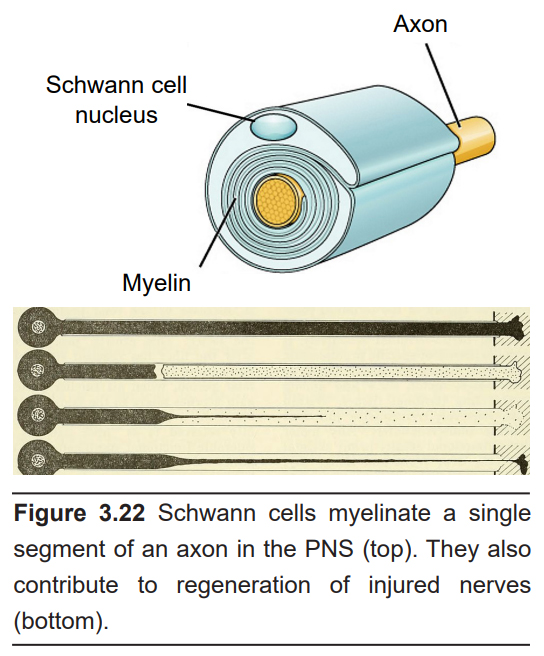
- Schwann cells
Schwann cells are named after the German biologist who first described these glial cells. He found that there are cells that are wrapped around the axons of nerve cells that project towards muscles – myelin. These Schwann cells can only be found in the PNS.
The main action of Schwann cells is to provide a section of myelin sheath for PNS neurons, and in this way, they function similarly to the oligodendrocytes. Schwann cells produce only a single section of myelin, compared to oligodendrocytes, which myelinate multiple sections.
Schwann cells also function in the regeneration of injured axons. When nerves in the PNS are damaged after trauma, Schwann cells rapidly mobilize to the site of injury. Here, the remaining loose myelin hinders the regeneration process, and the Schwann cells destroy the extra cellular membranes. Schwann cells also produce signaling molecules that guide the injured axons to the correct targets, which helps the axon regrow.
- Microglia
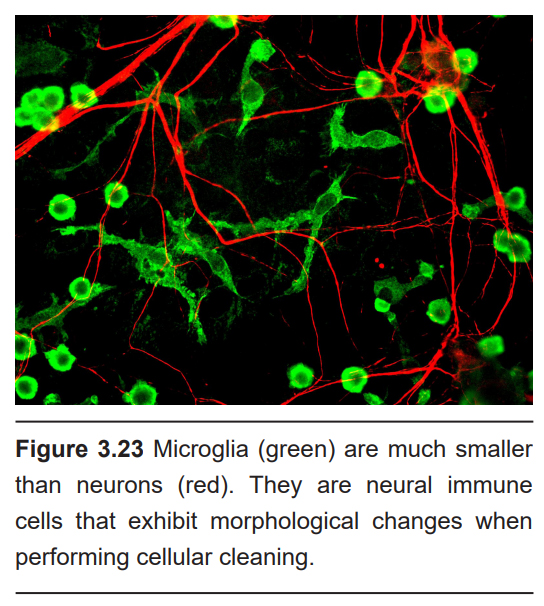
Microglia are a bit different from the other glial cell populations. For one, microglia are more immune cells rather than neural. They act as cellular scavengers that travel throughout the brain and spinal cord. It is estimated that microglia make up 10-15% of all cells in the brain.
As immune cells, microglia identify and destroy clumps of proteins, dead / dying cells, or foreign pathogens that enter into the brain. After an injury to the CNS, like a traumatic blow to the head, microglia rapidly react to the area of the insult. The marker Iba1 is often used to identify when microglia are reacting to an injury.
- Ependymal cells
Along the inside of the ventricles are a lining of glia called the ependymal cells. These ependymal cells are columnar with small finger- like extensions called cilia that extend into the ventricles and into the central canal that runs down the inside of the spinal cord.
Ependymal cells produce CSF. In total, the body can make about half a liter of CSF each day (a little more than two cups.) The ependymal cells are part of a structure called the choroid plexus, the network of blood vessels and cells that form a boundary between the blood and the CSF.
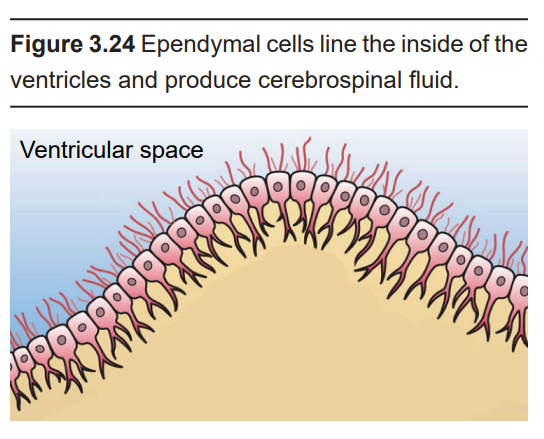
Image credits
Cover: Dana H. Simmons, Ph.D. dana-simmons.com
- 1 https://upload.wikimedia.org/wikipedia/commons/6/6d/GolgiStainedPyramidalCell.jpg https://upload.wikimedia.org/wikipedia/commons/1/1a/Human_hippocampus_2.5x.jpg
- 2 https://upload.wikimedia.org/wikipedia/commons/5/57/Camillo_Golgi_nobel.jpg https://upload.wikimedia.org/wikipedia/commons/3/30/Cajal-Restored.jpg
- 3 https://upload.wikimedia.org/wikipedia/commons/5/5f/Cell_membrane_detailed_diagram_simplified_ca.svg modified by Austin Lim
- 4 https://svgsilh.com/9e9e9e/image/2040692.html modified by Austin Lim
- 5 https://pixabay.com/photos/axe-old-lumberjack-blade-1008981/
- 6 https://upload.wikimedia.org/wikipedia/commons/0/0a/Anatomy_of_a_Neuron_with_Synapse.png modified by Austin Lim
- 7 https://upload.wikimedia.org/wikipedia/commons/7/74/Dendritic_arborization_patterns_2.png https://upload.wikimedia.org/wikipedia/commons/9/90/Submerged_Albizia_Saman_in_the_Mekong_ at_sunset_%28close-up_view%29.jpg
- 8 https://upload.wikimedia.org/wikipedia/commons/6/6e/Geometric_characteristics_of_dendritic_spines.png modified by Austin Lim
- 9 https://upload.wikimedia.org/wikipedia/commons/f/fb/MRNA-interaction.png modified by Austin Lim
- 11 Image courtesy of Connor Maltby
- 12 https://upload.wikimedia.org/wikipedia/commons/8/86/201704_microtubule.svg modified by Austin Lim
- 15 https://upload.wikimedia.org/wikipedia/commons/2/2a/Multiple_Sclerosis.png modified by Austin Lim
- 16 https://upload.wikimedia.org/wikipedia/commons/9/92/Neurons_uni_bi_multi_pseudouni.svg modified by Austin Lim
- 17 https://upload.wikimedia.org/wikipedia/commons/6/6c/Synapse_neuro-neuronale.png
- 3.19 https://upload.wikimedia.org/wikipedia/commons/5/56/Human_astrocyte.png https://upload.wikimedia.org/wikipedia/commons/3/34/Astrocytes_Wrapping_around_Blood_Vessel. png
- 21 https://upload.wikimedia.org/wikipedia/commons/2/2e/Oligodendrocyte_illustration.png
- 22 https://upload.wikimedia.org/wikipedia/commons/5/52/CeluladeSchwann.jpg https://upload.wikimedia.org/wikipedia/commons/0/03/Analysis_of_ development_%281955%29_%2817549733973%29.jpg modified by Austin Lim
- 23 https://upload.wikimedia.org/wikipedia/commons/0/0b/Microglia_and_neurons.jpg
- 24 https://upload.wikimedia.org/wikipedia/commons/c/ca/Glial_Cell_Types.png modified by Austin Lim
The Open Neuroscience Initiative is funded by a grant from the Vincentian Endowment Fund of DePaul University.
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License.
