10 Chapter 10: The Motor System
Cover Image
Alexander Rajan, PhD
Austin Lim, PhD (DePaul University)
Andrea Gaede, PhD
Editor: Katie Willis, PhD (University of Oklahoma)
The motor system refers to the nerve cells that are used to control our body. The two key roles of the motor system are to plan, control, and execute voluntary (deliberate) movements, and to control involuntary (subconscious or automatic) functions, such as digesting food.
The motor system is sometimes described as a top-down process: in a voluntary movement, neural activity in the frontal lobe sends commands down to motor neurons located in the brainstem or spinal cord, which in turn activate muscle groups.
In reality, motor control is more of a loop, rapidly communicating between the sensory cortex and motor cortex. Sensory information about limb position, posture, and objects in contact with the skin inform the descending motor plan. Simultaneously, the motor plan provides predictions about upcoming movement. Without knowing where you are, it’s difficult to plan a route to your destination, and getting feedback along the way helps your brain know if it needs to adjust the plan.
Accordingly, think about motor control as a set of nested loops, where motor and sensory processes are closely intertwined. The main loop is the descending (from brain to muscle) motor command and the ascending (from muscle to brain) sensory feedback. Examples of nested loops within this circuit could include spinal cord-mediated reflexes and communication between areas within the brain.
We will describe the process of motor control by first describing the signals that originate in the brain, then tracing that signal down through the brain stem, spinal cord, the neuromuscular junction, and finally the muscles.
Chapter 10 outline
10.1 Motor Control in the Brain
10.2 Modifiers of Descending Information
10.3 The Spinal Cord
10.4 The Muscles
10.1 Motor Control in the Brain
All voluntary (or non-reflexive) movements begin as signals in the brain. Specifically, the neurons involved in motor control are primarily found in the frontal lobe of the telencephalon, which includes areas such as primary motor cortex (or M1), premotor cortex, and supplemental motor areas. The posterior parietal cortex also contributes to movement.
Through this section, we will walk through the brain processes leading to voluntary motor action, beginning at the highest areas of the hierarchy.
Association cortices
Neural control of voluntary movement begins with high order thought processes which are carried out by two major associative areas, the prefrontal cortex and the posterior parietal cortex. These areas do not specifically correlate with specific muscle groups, and activation of these neurons do not necessarily cause muscle activity. Instead, these structures are important for initiation of motor control. Primates have many cortical areas dedicated to movement, which allows for fine control of small muscle groups, complicated patterns of movements, and long- term planning of motor action.
With respect to motor control, the prefrontal cortex (PFC) initiates the long-term planning or cognitive aspects of movements. For example, consider the motor actions related to brushing your teeth. PFC signals are more akin to “brushing is good for my hygiene and health”, rather than “move my arm and open my mouth.”
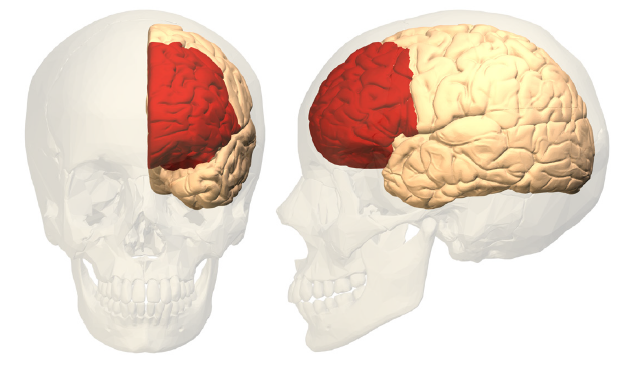
Figure 10.1 Anterior view (left) and lateral view (right) of one hemisphere showing the location of the prefrontal cortex (red).
PFC also helps determine if some motor action is appropriate for the specific situation. Think of a behavioral test where you are given two clickers, one to hold in each hand. You are told that when the experimenter shows you a green item, you should click with the right button. After repeating this behavior multiple times, you are told to switch – now, when you see a green item, you need to click the left button. In this experiment, PFC is responsible for deciding which motor pattern (left button or right button press) is appropriate in response to the stimulus.
PFC also works to weigh the consequences of motor actions and makes updates about future motor actions in similar or different circumstances. The exact same motor action produces different results depending on the specific situation, and PFC contributes to evaluating and predicting outcomes. For example, the Wisconsin Card Sorting Task is a human behavioral test that is strongly dependent on PFC activity. In this task, a patient is asked to classify cards based on criteria, such as number, shape, or color. The patient is told whether they are correct or wrong; however, the criteria will occasionally change without them being explicitly informed. The ability to shift to a new set of rules is PFC mediated.
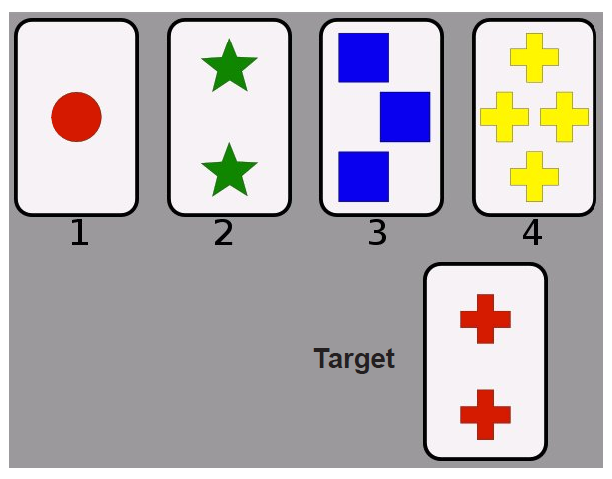
Figure 10.2 In the Wisconsin Card Sorting Task, the target card (bottom) could be placed in category 1 (matching color), category 2 (matching number of objects), or category 4 (matching shape). The ability to switch to a new set of rules uses prefrontal cortex.
The other major associative cortex contributing to motor function is the posterior parietal cortex, which is largely concerned with integrating somatosensory with visual information and determining an appropriate motor action. For example, if you were planning to get up from your seat to walk across the room, the posterior parietal cortex would take in the somatosensory proprioceptive information about how your body is positioned, and the visual information from the objects in the room to avoid running into them (recall the dorsal stream pathway; chapter 7.4).
Motor cortex
The motor cortex is made up of three closely related brain structures that contribute to execution of movement: premotor area (PM), supplemental motor area (SMA), and primary motor cortex (or M1). These motor cortex areas are found in the posterior aspect of the frontal lobe, directly adjacent to the central sulcus. The PM is the anterior most structure, and SMA is more dorsal to the PM along the medial aspect of the brain. M1 is posterior to that, bordering the central sulcus. Generally speaking, information from the associative cortices travel through PM, then M1, before projecting down through the brain stem and spinal cord.
These three structures are not an exhaustive list of motor cortex structures, however. For example, the frontal eye field (FEF) communicates with the extraocular muscles and mediates saccadic eye movements (chapter 7.4). The inferior frontal gyrus, or Broca’s area, contributes to motor processes related to language (chapter 14).
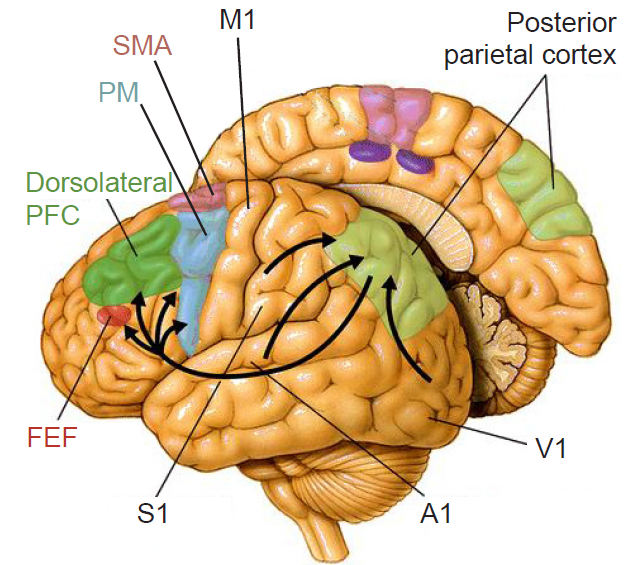
Figure 10.3 The posterior parietal cortex is part of the dorsal stream and integrates sensory information with motor commands.
The premotor area (PM) modulates motor output and generally activates prior to motor activity. For example, if monkeys are trained to press on a button on a delay in response to a specific light cue, PM neurons will increase activity upon presentation to the light cue, even before the arm or hand begins to move. Also, activity of PM increases as we imagine performing a complex series of finger patterns, such as during mental rehearsal of guitar or violin.
There is debate about the existence of a special population of cells in PM called mirror neurons. These are cells that are active during a movement, but also when that same movement is observed in another animal or person. Proponents argue that these neurons are involved in learning behaviors and for understanding the behaviors of others. However, given the complexity of the task of understanding others, it is unlikely to be encoded at the level of individual neurons.
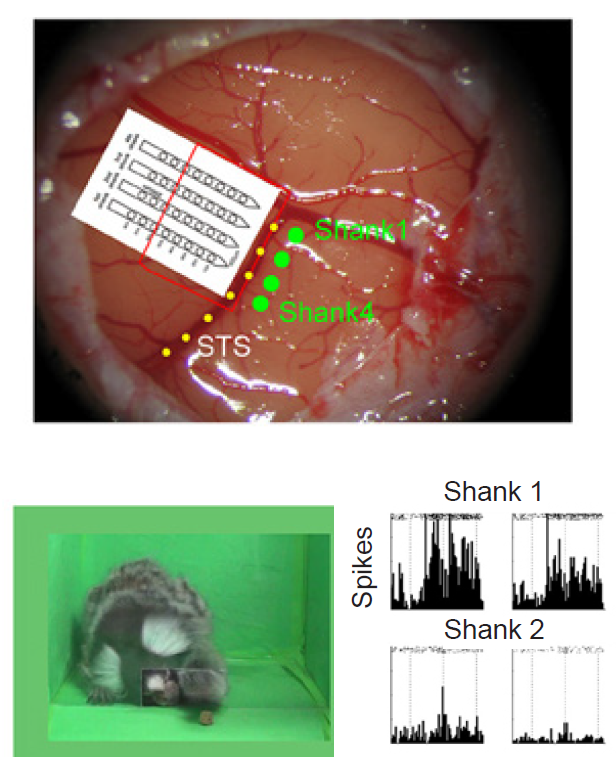
Figure 10.4 Surgically implanted electrodes (top) in a marmoset brain. While viewing a fellow marmoset reaching (bottom left), mirror neuron activity increases (bottom right).
The supplemental motor area (SMA) is upstream of primary motor cortex, but also sends downward projections through spinal cord. SMA communicates bilaterally, and lesioning this area causes deficits in manual coordination tasks that require both hemispheres to communicate. For example, in one monkey study, animals were presented with a table in which was a hole stuffed with a food reward. In order to obtain the food reward, the monkey needed to use one hand to push, and the other hand to catch. Lesions of SMA caused the monkey to try to push the food from both sides simultaneously. This suggests that SMA is responsible for the proper communication between motor commands in both hemispheres.
Primary motor cortex (M1) is a major motor control center, required for deliberate, voluntary movements, movements made in response to a “command.” Furthermore, motor cortex cells influence motor neurons, neurons that communicate down into spinal cord that, ultimately, influence muscles or glands. This connection is so strong that the motor cortex cells are sometimes called upper motor neurons. In this terminology, a lower motor neuron is found at the brain stem or spinal cord, and fires whenever the upper motor neuron sends a signal.
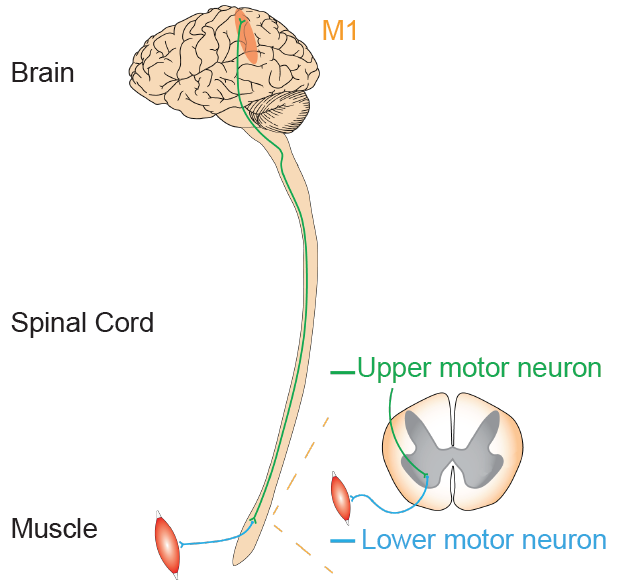
Figure 10.5 Relationship between upper motor neurons in M1 and the lower motor neurons further down of the signaling pathway.
In the 1930s, neurosurgeon Wilder Penfield conducted several brain operations to treat patients with severe epilepsy. However, since the brain has no pain receptors, he was able to remove a portion of the skull under local anesthesia while the patients were awake and responding to his questions. During surgery, the goal was to electrically stimulate portions of the cortex to determine the origin of the seizures, and to ensure that areas critical to speech and hand movement are left untouched so that the patient will not have major impairments following surgery. This is currently done in neuro-oncology to reduce the loss of critical motor function and overall morbidity.
Penfield progressively moved across different brain areas of M1 while using an electrode to stimulate patches of the cortex. He had two major observations. First, stimulation caused contralateral activity: that is, stimulating the left side of the brain affects the muscle activity of the right side of the body. Secondly, by systematically moving across M1, he observed that different populations of neurons are responsible for communicating with specific muscle groups. For instance, dorsal M1 activates hip and trunk muscles, while more lateral M1 activates muscles of the face.
Penfield discovered that within the motor cortex, different muscle groups are laid out in a rough topography, meaning that neurons that control the thumb are near other neurons that control the thumb, and near other populations of neurons that control the index finger. A graphical representation of this map is called the “motor homunculus,” in which body parts with larger representations in the brain are shown with larger size (much like the sensory homunculus; chapter 8.3).
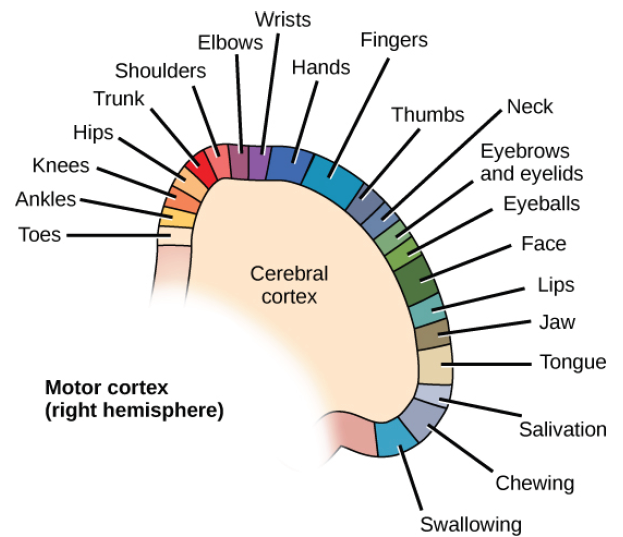

Figure 10.6 Topographical organization of the motor cortex (top), showing that neurons that control adjacent body parts are often adjacent themselves. The motor homunculus (bottom) is a representation of how much motor cortex is dedicated to control over a specific body part.
Clinical connection: Prosthetic limbs
If activation of specific neurons can produce muscle activity, then decoding these descending signals could be used to help amputees control their prosthetic limbs. This technology is called a brain- machine interface. In a technique called an electrocorticogram (eCoG), the surgeon puts a high-density electrode array capable of sensing neural activity directly onto the surface of the cortex of M1. With this strategy, it is possible to detect electrical activity at high spatial resolution (hundreds of microns) and temporal resolution (hundreds of microseconds), which is critical for movements as precise and rapid as muscle movement. ECoG grids have already been successfully implanted in people with tetraplegia, allowing them to control external devices such as a cursor on a screen or a prosthetic limb.
In the early 2000s, researchers discovered that complex preprogrammed actions, such as opening the mouth and bringing a hand to the mouth, could be activated from a single parcel of brain regardless of where the hand was located in space – on your knee, on the table in front of you, or to the side, for example. In these experiments, the researchers delivered long stimulation trains, similar in duration to that observed in large muscle group movement – approximately a half a second or longer. Further experiments developed a gestural map, and even more complex behaviors such as reaching, climbing, and defensive postures.
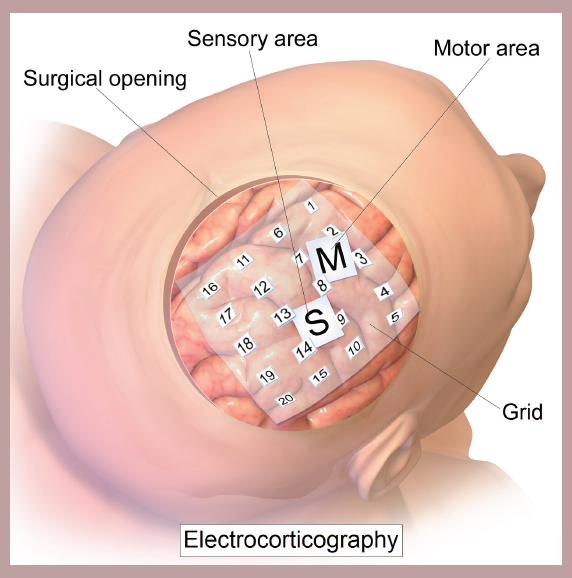
Figure 10.7 An eCoG allows a machine to read cortical electrical activity with high temporal and spatial precision, which can be used to control prosthetic limbs.
Sensory feedback
An important component of healthy movement is somatosensation, the process of sensing and perceiving the body (chapter 8.) One aspect is proprioception, the perception of our body in the world. For example, if you close your eyes, you can still feel if you are slouching and if your hands are in front of you or to your sides. Even if you were to move your arms forward, such as when reaching out to grab a nearby object, you could do it reasonably well without visual feedback. In order to make an accurate movement, the brain needs to know how well the actual movement matches the intended movement. If you are making an error, the sooner it is discovered, the sooner you can update your movement plan and adjust the movement.
Receptors in our muscles, tendons, joints, and skin detect contraction, stretch, and vibration. Sensory neurons carrying this information synapse onto neurons in the spinal cord, sending sensory information to both motor neurons and also up to the brain, particularly the primary somatosensory cortex (or S1; chapter 8). Here, several bidirectional connections are made onto neurons in the adjacent motor cortex. Through these signaling communication routes, sensory information is used to update the motor commands and correct movements that go awry.
10.2 Modifiers of Descending Information
Although the descending signals from M1 are the major motor command signals that regulate activity, that signal is fine-tuned through the action of two major brain structures, the cerebellum and a series of nuclei called the basal ganglia.
The Cerebellum
The cerebellum (“little brain” in Latin) is the most prominent structure of the hindbrain, located at the ventral-most part of the brain. The structure is evolutionarily ancient, and the general architecture and cell types of the cerebellum are conserved between teleost fish and mammals. The human cerebellum contains 69 billion neurons, which represents 80% of the total number of neurons in the human brain, despite being physically only 10% of the total brain mass.
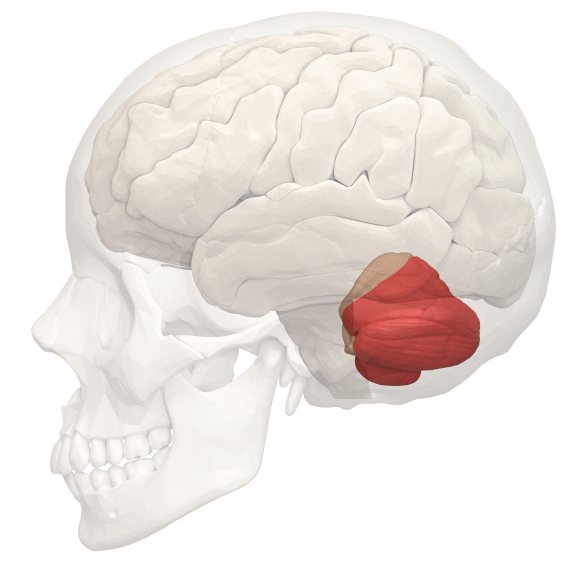
Figure 10.8 The cerebellum is relatively small, but contains more neurons than the rest of the brain.
Since the time of Galen in Ancient Greece (~200 CE), behavioral functions related to sensorimotor control have been ascribed to the cerebellum. In 1824, Marie-Jean-Pierre Flourens demonstrated that pigeons with cerebellar damage have poor wing flap coordination, resulting in a diminished ability to fly. Anatomist Jan Evangelista Purkynĕ (1787–1869) described the major output cell of the cerebellar cortex, and histological studies conducted by Santiago Ramon y Cajal (1909) later described cell morphology and the layers of the cerebellum.
The idea that the cerebellum is key for coordinated movement remains central to cerebellar research. The cerebellum plays a role in integrating sensory information to produce coordinated movement, refining motor-related outputs to learn motor tasks, and processing cognitive and executive functions.
Nonmotor functions of cerebellar circuits have not received significant attention until recent years. Stimulation or lesion of deep cerebellar nuclei produced various autonomic and complex behavioral outputs, and associative learning occurs in the cerebellum. Cerebellar abnormalities like Purkinje cell loss may be involved in autism. Furthermore, cerebellar cognitive affective syndrome (CCAS) is characterized by dysfunction in executive tasks (e.g. planning, working memory, abstract reasoning), impaired visual-spatial memory, changes to personality and emotional control, and problems with language production, demonstrating that cerebellum contributes to much more than just motor related behaviors.
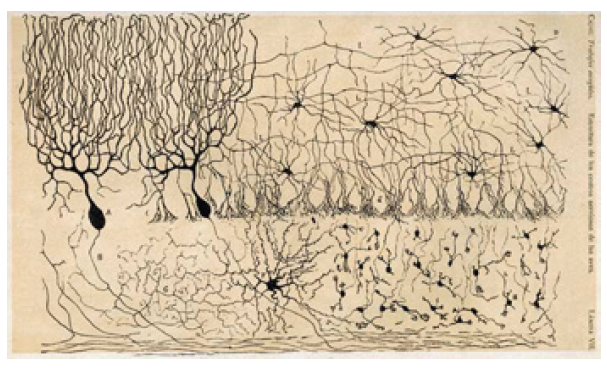
Figure 10.9 Early illustration by Ramon y Cajal of the different types of neurons found in the cerebellum.
Anatomy and function of the cerebellum
The cerebellum is composed of an external layer of gray matter (the cerebellar cortex), an internal core of white matter, and three pairs of deep nuclei (the fastigial nucleus, the interposed nuclei, and the dentate nucleus). The cerebellum is connected to the dorsal brain stem by three pairs of peduncles, or stalks of tissue: the inferior cerebellar peduncle, the middle cerebellar peduncle, and the superior cerebellar peduncle.
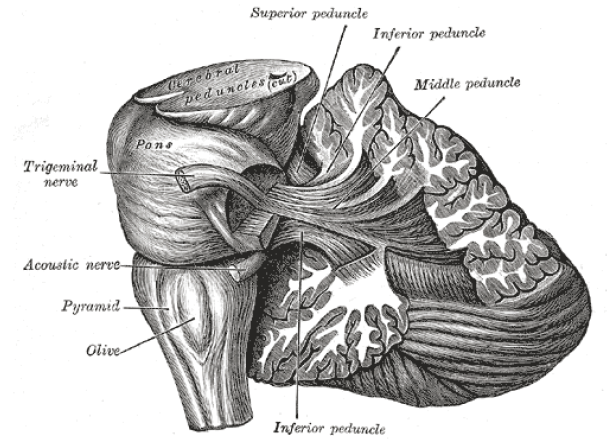
Figure 10.10 The cerebellum is attached to the brain stem via three large pairs of nerve fibers.
The cerebellum is organized in a series of regular, repeating units. It has a somatotopic organization with different regions of the cerebellum receiving afferent fibers from different sensory systems and projecting to different motor systems.
The surface of the cerebellum has a series of parallel folds called lobules. Two transverse fissures, the primary fissure and the posterolateral fissure, divide the cerebellum into three lobes. On the dorsal surface, the primary fissure separates the anterior and posterior lobes, while on the ventral surface, the posterolateral fissure separates the body of the cerebellum from the flocculonodular lobe. The midline region is called the vermis (Latin for “worm”), and on either side of the vermis is a cerebellar hemisphere.
Functionally, the cerebellum can be divided into three defined regions with roles in distinct types of movement.
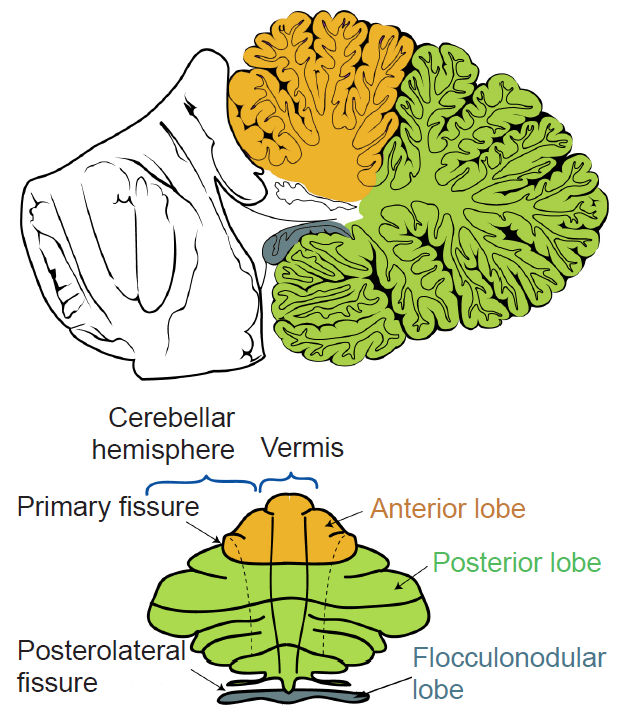 Figure 10.11 A midsagittal view of the cerebellum (top) and an “unrolled” view (bottom) labeling the major anatomical structures.
Figure 10.11 A midsagittal view of the cerebellum (top) and an “unrolled” view (bottom) labeling the major anatomical structures.
- The vestibulocerebellum, composed of the flocculonodular lobe, is the most evolutionarily-conserved region of the cerebellum. It integrates visual and vestibular inputs, relaying information about head and body position in space. It can be thought of as two components, a medial aspect and a lateral aspect.
Medial vestibulocerebellum controls muscles of the trunk and head, which regulate posture. It also communicates with the limb extensor muscles, which help maintain balance at rest and during locomotion.
Lateral vestibulocerebellum controls eye movements and coordinates head and eye movements. In clinical observations, patients with lesions in the lateral vestibulocerebellum display deficits in smooth pursuit eye movement towards the side with the lesion.
- The spinocerebellum, which includes the vermis and intermediate regions of the hemispheres, receives somatosensory and proprioceptive inputs from the spinal cord and is important for locomotion and extremity movement. The distal muscles of the limbs and digits are controlled by neurons in the intermediate regions of the cerebellar hemispheres that receive somatosensory inputs from the limb.
Somatosensory information (touch, pressure, proprioception; chapter 8) is passed to the spinocerebellum from the spinal cord. These connections provide feedback to the organism about its changing position and environment so that comparisons and adjustments can be made. Projections into the spinocerebellum display approximate somatotopy. There are two functionally distinct ascending pathways into spinocerebellum, both carrying information from spinal interneurons.
The dorsal spinocerebellar tract carries joint and muscle somatosensory information, providing the cerebellum with sensory feedback related to both voluntary and involuntary movement.
The ventral spinocerebellar tract is only active during voluntary movements. It sends a “copy” of spinal motor neuron activity to the cerebellum, informing the movement commands assembled at the spinal cord level.
Together, these spinocerebellar connections are important for motor learning. Sensory information from the actual movement (dorsal) is compared with the expected movement (ventral). If there is a mismatch, the motor command gets modification to achieve the desired output.
The cerebellum also initiates feed-forward control of muscle activity to regulate the timing of movements. This is anticipatory activity that causes muscle contractions to generate smooth and accurate motion.
- The cerebrocerebellum, composed of the lateral portions of the cerebellar hemispheres, is the most evolutionarily recent region, and is much larger in humans and apes than other basal-order mammals. As the name implies, these structures communicate with the cerebral cortex. Outputs of the cerebrocerebellum travel to motor and premotor cortices, and function in planning and executing movement. The cerebrocerebellum also targets nonmotor associative areas, such as prefrontal and posterior parietal cortex, contributing to cognitive functions. Neuroimaging and neuropsychological studies demonstrate cerebellar activation during tasks designed to evaluate attention, planning, working memory, abstract reasoning, language, pain, emotion and addiction.
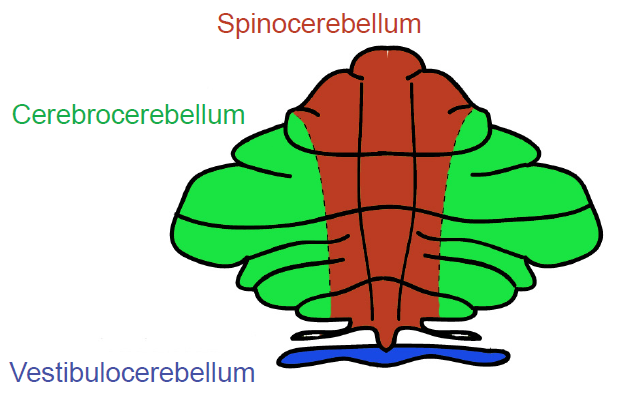
Figure 10.12 Functional divisions of the cerebellum.
Clinical connection: Ataxia
Ataxia is a condition characterized by poor coordination of voluntary movements, atypical eye movements, poor balance, or changes in gait. They may exhibit dysarthria, a speech disorder resulting in difficulty with articulating language. Acute alcohol intoxication produces an approximation of the symptoms of ataxia.
A wide variety of causes can result in ataxia, such as cerebellar injury from a stroke or tumor, toxin exposure, radiation poisoning, hereditary diseases, or diet-induced (gluten) autoimmune mediate cell death. Treatment of ataxia is most effective when the underlying cause is addressed, and ataxia may be completely reversible or untreatable. Medication and physical therapy may also help treat the symptoms.
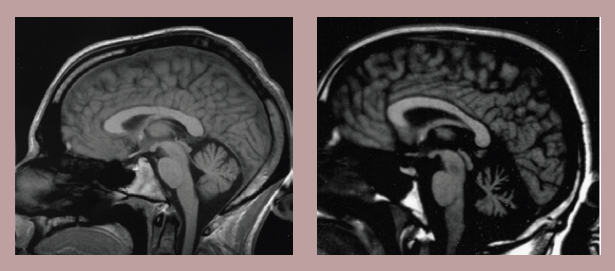
Figure 10.13 MRI showing a healthy cerebellum (left) and one experiencing degeneration associated with ataxia (right).
Cellular organization of the cerebellum
The cerebellar cortex is arranged into three layers with distinct populations of neurons.
The deepest layer, called the granular layer, is the input layer of the cerebellar cortex. It contains a substantial number of small, densely packed excitatory neurons called granule cells, which are the primary unit of this layer. The granular layer also includes more sparsely distributed inhibitory interneurons (Golgi cells, Lugaro cells and chandelier cells) as well as excitatory interneurons (unipolar brush cells). Mossy fibers, one of the two major types of afferent projections into the cerebellum, terminate in the granular layer. Mossy fibers form synaptic complexes with granule cells and Golgi cells called cerebellar glomeruli that allow neurotransmitter spillover and crosstalk.
The middle layer is called the Purkinje cell layer and constitutes the output layer of the cerebellar cortex. It consists of a single layer of Purkinje cells, which have giant cell bodies and a broad dendritic arbor that stems from a central dendrite and fans in a single plane, extending upward into the outer layer of the cerebellar cortex. Purkinje cell axons are responsible for the output of the cerebellar cortex, projecting to deep cerebellar nuclei in the white matter or vestibular nuclei in the brain stem where GABA is released.
The external layer of the cerebellar cortex is the molecular layer, which contains the apical dendrites of the Purkinje cells and parallel fibers, the axons of granule cells. These parallel fibers ascend from the granular layer to the molecular layer where they bifurcate to form branches that extend mediolaterally, parallel to the long axis of the folia/lobules, thus giving them their name.
Also in the molecular layer are two populations of inhibitory interneurons, the stellate cells and basket cells, which provide feed-forward inhibition to Purkinje cells.
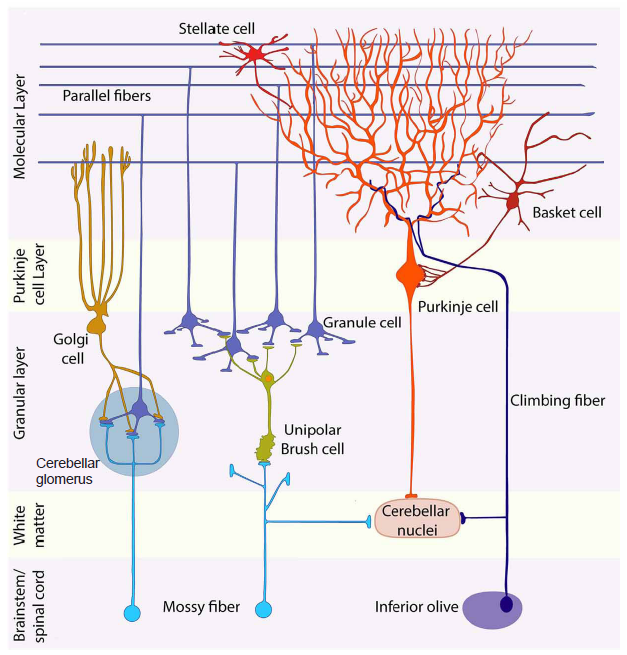
Figure 10.14 Cartoon illustrating the cellular organization of the neurons of the cerebellum.
The Basal Ganglia
The basal ganglia are a series of subcortical brain structures that are intimately involved with various aspects of movement, such as voluntary motor activity, habit learning, and the selection of actions. Despite being major structures involved in motor functions, none of the components of the basal ganglia directly send projections down the spinal cord. Rather, they communicate mostly within themselves before signaling through the motor cortex. While the connections between basal ganglia structures have been largely mapped out, the specific functions of each individual structure in isolation is not entirely clear.
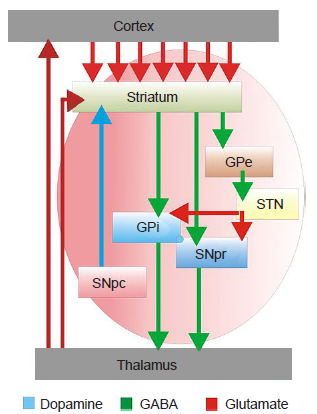
Figure 10.15 Diagram showing the connectivity and the nature of the signaling between different nuclei of the basal ganglia.
Diseases of the basal ganglia include movement disorders such as Parkinson’s disease, dystonia, Huntington’s disease, and Tourette’s, as well as complex psychiatric disorders including addiction and obsessive-compulsive disorder (OCD).
Striatum
The striatum is the largest of the basal ganglia structures. It is the main input site of the basal ganglia, with projections coming from both cortical and limbic structures such as thalamus and amygdala.
On a cellular level, the majority of neurons in the striatum are GABAergic cells called spiny projection neurons (SPNs), or medium spiny neurons (MSNs). These neurons express different subtypes of dopamine receptors, and some of them are excited by dopamine while others are inhibited. Mixed within the SPNs are several populations of interneurons with various electrophysiological properties that utilize different neurotransmitter systems, such as GABA, acetylcholine, and nitric oxide.
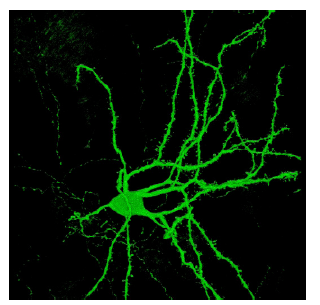
Figure 10.16 Confocal image of a striatal spiny projection neuron identified with green fluorescent protein (GFP).
The striatum is divided into two anatomically and functionally different structures, a dorsal and ventral striatum.
Dorsal Striatum
The dorsal striatum is made up of two components, the caudate nucleus and putamen. In rodents, the two are indistinguishable.
Functionally, dorsal striatum contributes to learning of habitual behaviors. Habits are learned motor patterns that can be performed without full attention and generally inflexible due to repeated training. Habits can either be adaptive (such as changing physical stance in response to different environmental threats) or maladaptive (such as compulsive hair pulling or in substance use disorder).
An example of a behavioral test of habit learning is a serial reaction time test, where one of four visual stimuli are presented and the patient is expected to make a motor response, such as pushing on a corresponding button. If the pattern of stimuli repeats with some regularity (button 2 is often followed by button 4, for example) patients can unconsciously learn the repetition of the pattern, decreasing reaction time.
The caudate is particularly important for goal-directed actions, which are sets of motor behaviors that are made in response to knowledge of which actions lead to which outcomes. such as the complex motor actions made while performing an operant conditioning task. An experimental paradigm for this behavior would be a Skinner box, where an animal lever- presses or nose-pokes for a food or sugar reward (chapter 11.2).

Figure 10.17 Skinner box can be used to measure operant conditioning, a dorsal striatum-mediated behavior.
The putamen contributes to motor-associated procedural learning tasks, like a mirror tracing task (chapter 13.1), which involves learning how to draw with a pencil while watching only a reflection of your hand. As with the serial reaction time test, improving at these behaviors happens unconsciously.
Anatomically, the dorsal striatum receives excitatory glutamatergic inputs from both motor cortex and thalamus. It also receives dopaminergic projections from another basal ganglia structure, the substantia nigra pars compacta, a communication route called the nigrostriatal pathway.
The only output of the dorsal striatum are the GABAergic spiny projection neurons (SPNs), which exist in two different types with two different targets. One population, the direct SPNs (dSPNs), express dopamine D1 receptors and send axons into the internal globus pallidus (GPi). When this direct pathway is activated, it increases motor activity. The other output neurons, the indirect SPNs (iSPNs), express dopamine D2 receptors and project into the external globus pallidus (GPe). Activation of this indirect pathway decreases motor activity.
For healthy behavioral output, the dSPNs drive the intended motor action, while the iSPNs shut down competing motor actions. The balance of these systems is closely regulated, and imbalance of this signaling (such as in Parkinson’s disease) leads to motor disturbances.
Clinical connection: Huntington’s Disease
Huntington’s disease is a rare neurodegenerative movement disorder resulting from various dysfunctional signaling pathways of the basal ganglia. Symptoms include hyperkinesia (uncontrolled movement), poor coordination, and cognitive and psychiatric changes eventually leading to dementia. The onset of symptoms happens when a person is in their 30s-50s, and prognosis is generally fatal within 15 years after diagnosis. There is currently no cure for the disease.
Huntington’s disease passed through families in an autosomal dominant manner. A protein called huntingtin (htt) is implicated, and mutant huntingtin has several repeats of the amino acid glutamine. The greater the number of repeats, the more severe the symptoms and the earlier the onset of the disease. A leading theory suggests that mutant htt accumulates inside neurons, leading to neurodegeneration.
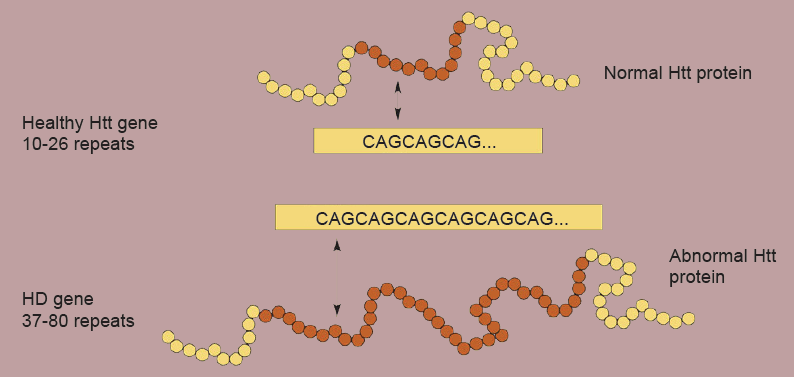
Figure 10.18 Longer CAG repeats in the Htt gene code for abnormal proteins. Greater number of repeats are associated with earlier age of onset of symptom presentation.
Ventral Striatum
The ventral striatum includes the olfactory tubercule and the nucleus accumbens, which is further subdivided into core and shell.
Importantly, these ventral striatum structures are important for reward, motivation, and aversion. As with dorsal striatum, dopamine signaling is critical for mediating these behaviors. Increased dopamine signaling in ventral striatum is a “learning signal” that encourages the organism to repeat that behavior again. Dopamine is released in response to unexpected rewarding stimuli, such as getting a sip of a sugary drink or engaging in sexual contact. Many drugs of abuse, such as nicotine and cocaine, artificially drive up this dopamine signal, which leads to repeated use and drug seeking (chapter 11).
The major inputs into the ventral striatum are glutamatergic afferents from prefrontal cortex and limbic areas, such as hippocampus, amygdala, and thalamus. The dopaminergic inputs come from the ventral tegmental area in the midbrain.
Globus Pallidus (GP)
In dissection, the GP appears as a pale globe. It is subdivided into two components with functionally different connections. The internal globus pallidus (GPi) is movement promoting, and receives inputs from the dSPNs of the dorsal striatum. On the other hand, the external globus pallidus (GPe) is movement inhibiting, and receives dorsal striatal afferents from iSPNs.
Subthalamic Nucleus (STN)
The STN is part of the indirect pathway, downstream of the GPe. The GPe makes up the axonal inputs into the STN, which are inhibitory GABAergic projections. The output of the STN are glutamatergic signals into the GPi.
In a medical intervention called deep brain stimulation (DBS), a surgeon implants permanently indwelling electrodes directly into brain tissue. These electrodes are controlled by an external battery pack that delivers pre- programmed stimulation protocols. DBS in the STN is used to alleviate the symptoms of Parkinson’s disease.
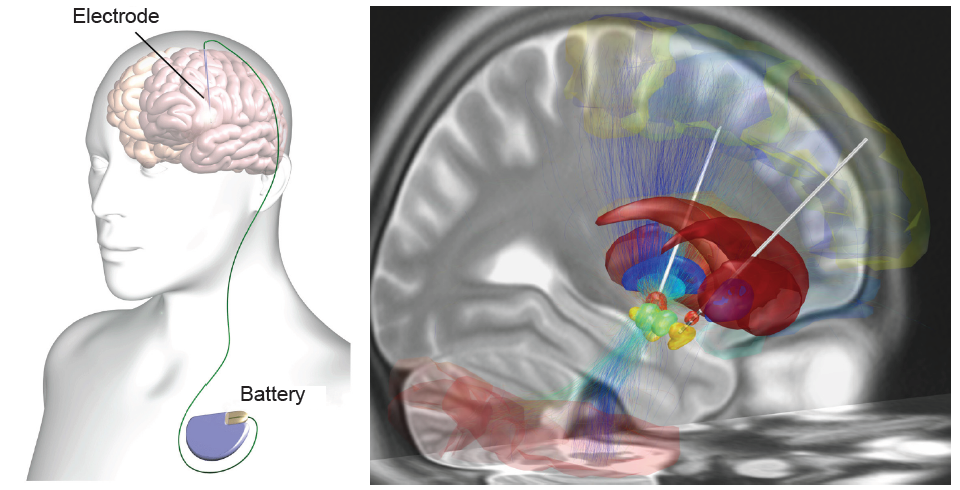
Figure 10.19 A deep brain stimulator (left) is controlled by an external battery. During surgery, the STN (right) can be targeted as a treatment for Parkinson’s disease.
Substantia Nigra
The SN is a midbrain structure that appears darker in dissection due to heavy expression of neuromelanin across cells in these areas. It is the largest midbrain structure.
SN is divided into two areas. The substantia nigra pars compacta (SNpc) contains several dopamine expressing neurons that project into the dorsal striatum. Of clinical significance, these neurons experience selective neurodegeneration in Parkinson’s disease.
The substantia nigra pars reticulata (SNpr) is an anatomically distinct area of the SN. It receives GABA-ergic inputs from the dorsal striatum and excitatory inputs from STN. The major output of SNpr is GABAergic; these axons terminate in the thalamus.
Clinical connection: Parkinson’s Disease
Parkinson’s disease is a neurodegenerative movement disorder that causes bradykinesia (slowness of movement), a resting tremor, muscle rigidity, and changes to posture and locomotion. Although most symptoms are motor, there are mild cognitive and psychiatric changes, such as apathy, anhedonia, mood disturbances, or depression. Advanced age is the primary risk factor, as an estimated 1% of people over the age of
60 develop PD. Other environmental factors contribute to risk, such as repeated traumatic brain injury (suspected in Muhammad Ali) or occupational exposure to heavy metals, insecticides, or other neurotoxins. A small percentage of cases are early onset (21 – 50 years old; Michael J. Fox was diagnosed at 30) and have a strong genetic component. The disease causes significant decreases in life expectancy and quality of life.
A loss of dopaminergic neurons of the SNpc contributes to basal ganglia circuitry disruption. When less dopamine is released into dorsal striatum, the dSPNs are less active while the iSPNs are more active. This imbalance in the output leads to the motor symptoms of the disease. However, the cause of PD is still unknown; the formation of intracellular protein aggregates called Lewy bodies and oxidative stress leading to neurodegeneration are among some of the proposed mechanisms.
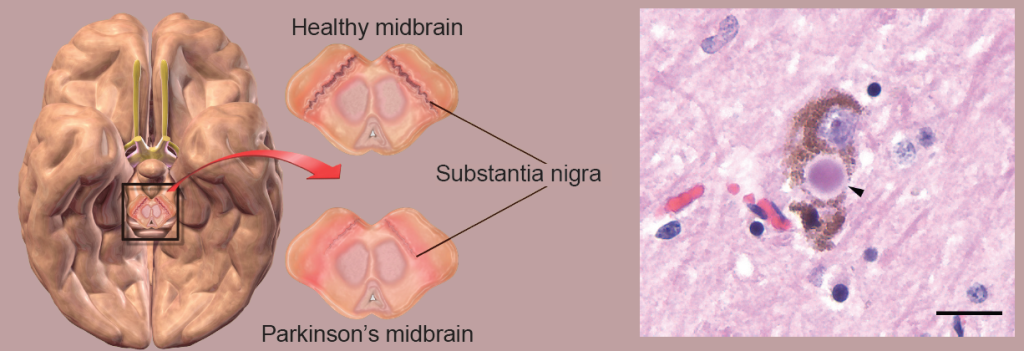
Figure 10.20 The substantia nigra selectively degrades in PD (left), possibly due to accumulation of Lewy bodies inside the neurons (right, arrow head).
10.3 The Spinal Cord
Spinal cord anatomy
The spinal cord extends posteriorly from the brain stem, surrounded by the vertebrae. In a typical human, the spinal cord is about 1 cm in diameter. Generally, the diameter of the spinal cord decreases from anterior to posterior, with two exceptions: an enlargement in the mid-cervical region to accommodate the extra nerves of the arms and hands, and a smaller enlargement in the lumbar region to accommodate extra nerves of the legs and feet (chapter 2.1).
The output neurons of the motor cortex carry voluntary motor information and send their axons down through two major descending communication routes. At the level of the brain stem, both pass through the medullary pyramids, paired anatomical structures found on the ventral surface of the medulla. Because of this, both descending axonal tracts are also called the pyramidal tracts.
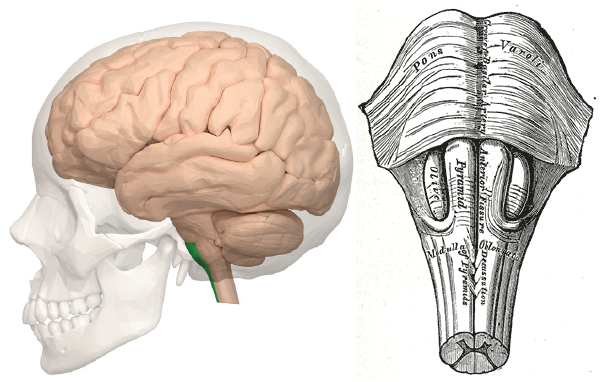
Figure 10.21 Position of the pyramids in the ventral spinal cord (left, green) and its location along the midline (right).
The corticobulbar tract is made up of axons that terminate in various brain stem motor nuclei. These nuclei communicate directly with several cranial nerves, particularly those with motor function: The trigeminal nucleus (CN V) modulates muscles of chewing, the facial nucleus (CN VII) regulates facial expressions, the glossopharyngeal nucleus (CN IX) controls the muscles involved in speech and swallowing, and the hypoglossal nucleus (CN XII) controls the muscles of the tongue.
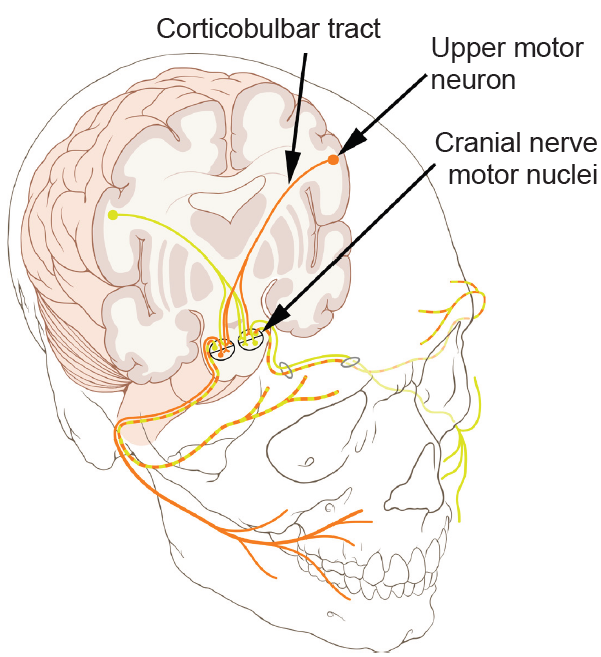
Figure 10.22 The corticobulbar tract originates in M1 and projects into the brain stem motor cranial nerve nuclei that control facial muscles.
The other major output of M1 are the upper motor neurons whose axons are the corticospinal tract, which runs down the spinal cord through white matter. This pathway has two different communication components.
Upwards of 90% of the corticospinal tract are axons of the lateral corticospinal tract, which sends voluntary motor control to the contralateral distal limbs, such as in the arms, hands, legs, and feet. Some neurons of M1 are Betz cells, the largest known neurons in the brain. Most of this tract crosses hemispheres at the pyramidal decussation, runs down the contralateral spinal cord, then forms synapses onto the lower motor neurons in the ventral horn. These lower motor neurons then send nerve rootlets out of the ventral surface of the spinal cord, then form a neuromuscular junction with the muscles.
The remaining output is the anterior corticospinal tract, which carries information to the muscles of the trunk, such as our shoulders or pectorals. This signaling from the brain stem descends ipsilaterally, but eventually decussates at the spinal cord, resulting in contralateral control. Here, as above, the neuron forms synapses with the lower motor neurons found in the ventral horn.
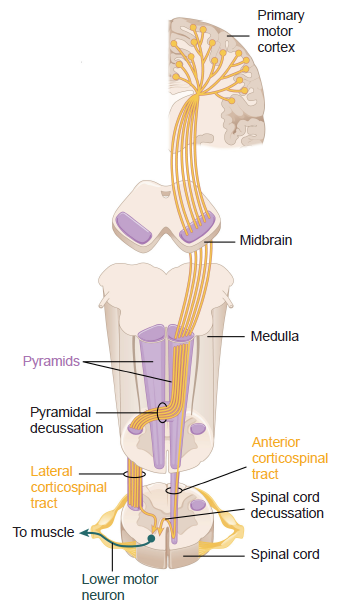
Figure 10.23 Connectivity of descending motor control pathways
Reflexes
Reflexes are involuntary motor responses that are performed automatically and independent of brain signals (although some can be suppressed voluntarily, with extra effort). Reflexes involve very simple circuits, sometimes consisting of only two populations of neurons: Sensory information comes in from the periphery, synapsing onto motor neurons in the brainstem or spinal cord. Reflexes can take place as quickly as 1/100th of a second!
A simple reflex is the patellar reflex, or the “knee-jerk” or myotatic reflex (described in more depth in chapter 8; figure 8.27). The stretching of the tendon causes the muscle to elongate, sensory receptors detect this change in muscle length, and the corresponding signal is sent into the spinal cord. This sensory neuron synapses directly onto a motoneuron, which then tells the thigh muscle to contract, making the kicking “knee-jerk” movement. Since only one synapse is involved in producing this response, this is an example of a monosynaptic reflex arc.
The reflexive kick is controlled at the level of the spinal cord and cannot be intentionally suppressed by descending motor pathways no matter how hard you concentrate.
Central pattern generators (CPG)
Central pattern generators (CPGs) are networks of cells that are capable of producing intrinsic motor responses even in the absence of sensory or brain inputs. These motor responses are usually well-rehearsed, repetitive, and happen at the unconscious level, meaning that certain behavioral outputs can be performed independently of signals upstream in the motor cortex. Some example motor responses driven by CPGs include diaphragm movement (respiration), alternating leg swinging and foot flexing (walking), and the progressive contraction of up to 25 pairs of muscles in the tongue and mouth (swallowing). Some CPGs are located in the brain stem (respiration) and others throughout the spinal cord (locomotion).
CPGs have been observed in several organisms, ranging from insects, crayfish, birds, and mammals including humans, hinting that their evolution was highly adaptive for survival. Imagine how difficult it would be to survive if every breath required a conscious thought!
The CPG is not a standalone driver of motor activity, however. The circuits receive signals from higher brain areas which can modify their characteristics. Imagine that you want to consciously hold your breath, or intentionally walk in some unusual or goofy way. In both cases, the descending signals from the brain are able to override the output motor command pattern that the CPG normally produces. Once you stop intentionally changing your motor behavior, the muscles return to their normal activity in response to CPG output.
CPGs of respiration
Getting oxygen is one of the most essential functions that an animal needs to perform for survival. As such, involuntary control of respiratory muscles through CPG activation is likely an evolutionary older mechanism. Respiratory control circuits are found at the level of the brain stem, specifically, the medulla and pons. One suspected CPG is a population of medullary cells called the pre-Bötzinger complex, which contains cells that change their activity in a cyclic pattern. Neurons in these areas are also sensitive to changes in blood chemistry (Chapter 9.3).
The major nerves involved with respiration are the phrenic nerve, which innervates the diaphragm, and the intercostal nerves, which predictably innervate the intercostal muscles. When we inhale (inspiration), the diaphragm contracts and the intercostals move the rib cage to increase the volume of the chest cavity, which allows the lungs to expand. During exhalation, the reverse process is true. These nerves receive innervation from neurons of the CPGs in the respiratory centers of the hindbrain (medulla and pons, specifically.)
The activity of the respiratory CPGs is so potent, that they can sometimes function while unconscious. Some deficits in respiratory CPGs are believed to be one cause of central sleep apnea, a disorder where a person stops breathing while asleep (chapter 12).
CPGs of locomotion
It is much more complex to describe the action of the muscle groups needed for a simple behavior of just walking down the sidewalk: the ankle dorsiflexors, the hamstrings, hip flexors, gluteus maximus and many more function at very precise moments, allowing us to propel ourselves forward. Importantly, this pattern of activity is rhythmic, and repeats at the frequency of stepping.
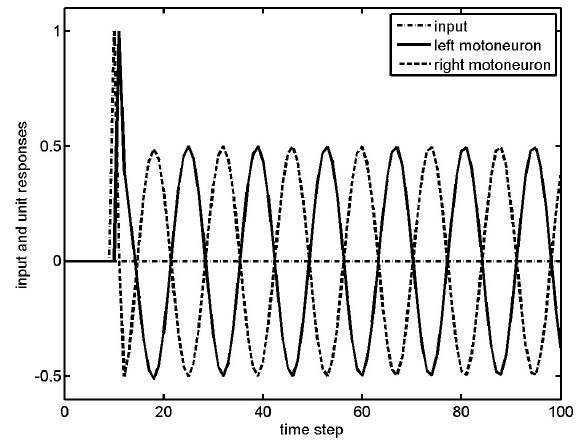
Figure 10.24 Locomotor CPGs produce alternating cyclic activity of motor neurons, which simplify walking.
Many earlier studies of spinal cord CPGs were conducted in different animal groups with unique locomotor patterns. Mollusks have crawling activity that is rhythmic, fish swim by sending alternative patterns of muscle activity through their body, and birds fly by rhythmic flapping of their wings.
In mammals, spinal CPGs were demonstrated in cats with a thoracic-level spinal cord transection. Following this surgery, the signals from the brain were unable to be communicated down to the hind legs, eliminating voluntary movement. They exhibited weakened muscular power, but were still able to stand for short times. If their front paws were put onto a stable platform while their hindlegs were put on a moving treadmill, the cat would involuntarily walk to keep up.
Humans also have CPGs for movement. One remarkable study looked at 37-year-old man who had injured his spinal cord in a football accident as a young man. His injury was at the level of C5, resulting in complete paralysis and a loss of sensation from the neck down. Over the following years, he gradually recovered a small amount of function. Fine manipulation skills and bladder bowel control never recovered, and he had not walked unassisted for ten years, experiencing tremendous weakness after taking a few steps.
Just days after beginning a physical therapy regimen consisting of assisted upright walking, upon lying down, the man reported alternating muscle flexion and relaxation of the hips, knees, and ankles similar to the pattern of activity seen in locomotion. He was unable to voluntarily stop these motions and could only get some rest by turning to the side.
This muscle activity was smooth and rhythmic, as would be seen in a healthy person walking.
Figure 10.25 Six-legged animals like insects have central pattern generators that simplify locomotion through coordinated neural activity that regulates muscle movement.
10.4 The Muscles
Neuromuscular junction (NMJ)
The signal from lower motor neurons communicates with the muscle at the neuromuscular junction (NMJ). The NMJ is similar to other chemical synapses; however, the postsynaptic cell is a muscle cell separated by about 30 nm. The postsynaptic site is the sarcolemma, the cell membrane of muscle cells, which has several folds to increase surface area.
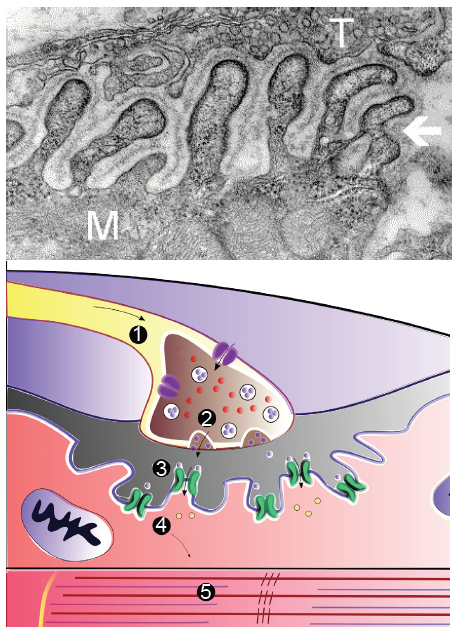
Figure 10.26 Electron micrograph image of an NMJ (top; T = axon terminal, M = muscle cell). Cartoon depicting the phases of signaling at an NMJ (bottom). The action potential (1) allows release of aceylcholine (2), activating nAChRs (3), allowing depolarization of muscle cells (4) and the contraction (5).
The axons of the lower motor neurons synthesize and release acetylcholine. Densely expressed in the sarcolemma are nicotinic acetylcholine receptors (nAChRs), ionotropic receptors that allow sodium influx and subsequent muscle cell depolarization upon acetylcholine binding. This depolarization results in muscle cell contraction.
Muscle anatomy
As mentioned above, motor functions can either be voluntary (moving your arm above your head) or involuntary (muscle contraction that leads to bowel motility, or heart beating). Although the nervous system influences both types of muscle activity, most of our discussion revolves around deliberate, voluntary skeletal muscle movement. The main action of muscles is to contract, a physical change of their shape where they become wider and shorter. For example, as you flex your arm, your bicep changes from long and thin to short and thick.
Different muscles have different characteristics driven by their shape and composition. For example, some muscles can be active for a long time without getting tired (maintaining your posture in your chair as you read this) while others can exert a lot of force but get tired quickly (lifting weights in the gym).
In studying muscles, some of the key measurements are where they connect to the bone, how long they are, how thick they are, and what type of muscle fibers they are made of. These measurements can be used to calculate how much force a muscle can generate and how quickly the joint can move.
Clinical connection: Myasthenia gravis (MG)
Myasthenia gravis (MG) is an autoimmune disorder characterized by muscle weakness, resulting in difficulty with speech, trouble with movement and swallowing, drooping eyelids, and double vision. Each year, an estimated 20 out of a million people get diagnosed with MG.
The muscle weakness seen in MG results from immune system-mediated destruction of the nAChRs expressed at the NMJ. Thus, when the lower motor neuron releases ACh, the muscle cells are unable to detect this release, so they fail to contract appropriately.
One therapeutic strategy involves inhibition of acetylcholinesterase, the enzyme that degrades ACh. This causes the synaptic ACh to remain in the synapse longer, increasing the chance that receptors get activated. Alternatively, autoimmune diseases like MG can be improved with immunosuppressant therapy. With successful treatment, MG usually does not result in changes in lifespan.

Figure 10.27 A patient with MG may show weakness of facial muscles.
Individual muscles, such as your biceps brachii, are made up of several muscle fascicles, which in turn are made up of muscle fibers. Muscle fibers are the individual cells in which contraction occurs. The functional units for contraction are called sarcomeres. It is the aggregated activity of hundreds of thousands of sarcomeres within each muscle fiber that generates the force with which you walk your dog or chew your food.
The contraction within sarcomeres happens between two proteins actin and myosin. Actin are the thin filaments that form the scaffolding, and myosin are thick filaments that pull the actin together, shortening the sarcomere and contracting the muscle. Although each sarcomere is small (1.5 to 3.5 µm), many are stacked back to back along the length of each fiber, pulling against each other to contract through the whole range of your joints. Many sarcomeres bundled in parallel provide the combined force that give muscles strength. The strength in all of your muscles comes from these tiny threads pulling against each other.

Figure 10.28 In a sarcomere, myosin (purple) and actin (red) overlap when the muscle flexes.
Muscle types
There are two key characteristics of muscles: their structure (striated due to the presence of sarcomeres or smooth), and whether they can be voluntarily controlled. Based on these characteristics, there are three types of muscles in humans and other vertebrates.
| Striated | Smooth | |
| Voluntary | Skeletal | |
| Involuntary | Cardiac | Smooth |
Skeletal muscle is what most people think of when they think “muscle.” They attach to bones with tendons, and exert force on your skeleton to create movement and exert force on objects.
Skeletal muscles are also called “voluntary muscles” because they are the muscles that move when you choose to make a movement. They are voluntarily contracted (and in reflexes) to move your body by moving your skeleton. There are two types of skeletal muscles.
Fast twitch muscles generate a lot of force quickly, but also tire quickly. They are used mostly in high intensity exercise like lifting weights and sprinting. Slow twitch muscles generate less force, but can work for a long time. They are used in endurance exercise like jogging. Many exercises use a combination of both.
Muscles come in several different shapes, defined by the arrangement of their fibers and how they connect to tendons. The different shapes allow for different properties: some can change length quickly, others change shape less but are stronger. Another consideration for shape is the geometry of the joint they cover: for example, the pectoralis muscle reaches from your chest across your shoulder, and so the muscle is wide and flat where it connects to your sternum, and narrows to a point where it connects to your upper arm.
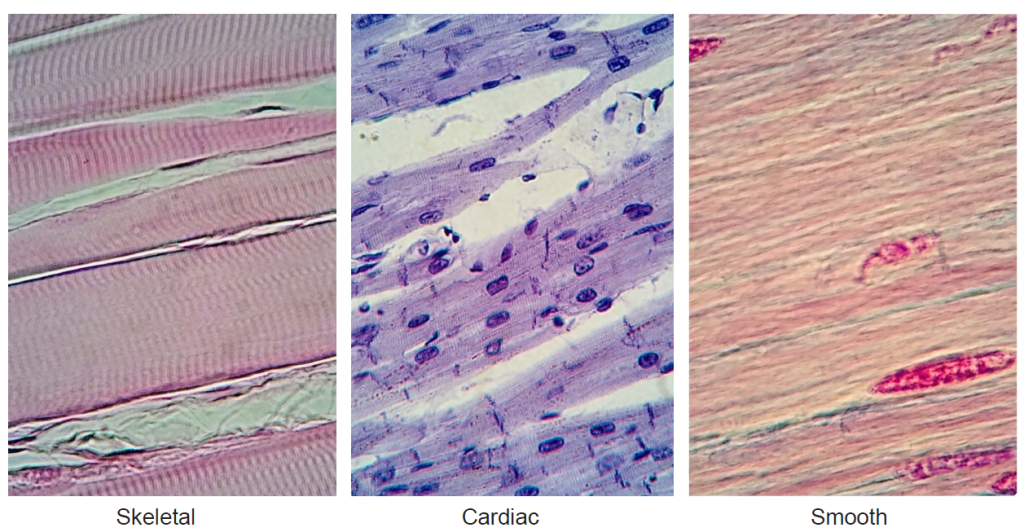
Figure 10.29 Different types of muscle as seen at 400x magnification. Muscle fibers are seen running from left to right. In skeletal and cardiac muscles, striations can be seen perpendicular to the muscle fibers.
Skeletal muscles work to move the body through a combination of cooperation and opposition. The agonist muscle is the main mover, like the biceps brachii when you flex at the elbow. Its movement is supported by synergistic muscles, including the brachialis and brachioradialis. These can also be fixators, providing stability and preventing or allowing rotation of the wrist while flexing at the elbow. Antagonist muscles are those that move in the opposite direction to the agonist. For the elbow, the antagonist is the triceps brachii, which lengthens during the bicep flex. Another example of an agonist-antagonist pair are the hamstrings to flex the leg, and the quadriceps to extend it.
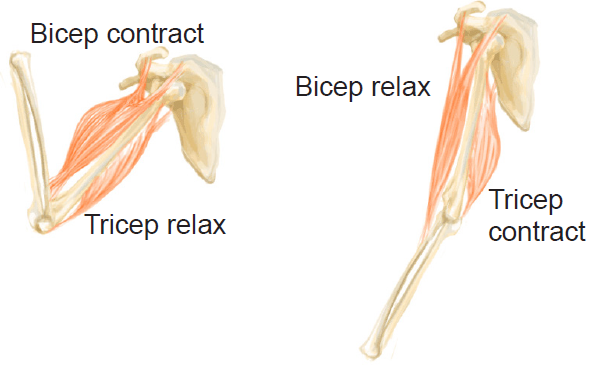 Figure 10.30 Skeletal muscles may be paired and produce opposing effects on a particular motion.
Figure 10.30 Skeletal muscles may be paired and produce opposing effects on a particular motion.
Smooth muscles are the muscles embedded within organs like your stomach and intestines, blood vessels, and bladder. They are also known as “involuntary” muscles because they are not under direct conscious control.
Cardiac muscle can be thought of as a hybrid between skeletal muscles and smooth muscles, in that they are striated like skeletal muscles but not under conscious control.
The Open Neuroscience Initiative is funded by a grant from the Vincentian Endowment Fund of DePaul University.
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License.
Image Credits
Cover: https://pixabay.com/photos/agility-ballet-dancing-athlete-1850711/
10.1 https://commons.wikimedia.org/wiki/File:Prefrontal_cortex_(left)_-_anterior_view.png https://commons.wikimedia.org/ wiki/File:Prefrontal_cortex_(left)_-_lateral_view.png
10.2 https://commons.wikimedia.org/wiki/File:WisconsinCardSort.png modified by Austin Lim
10.3 https://commons.wikimedia.org/wiki/File:Posterior_Parietal_Lobe.jpg
10.4 Suzuki W, Banno T, Miyakawa N, Abe H, Goda N and Ichinohe N (2015) Mirror Neurons in a New World Monkey, Common Marmoset. Front. Neurosci. 9:459. doi: 10.3389/fnins.2015.00459
10.5 https://commons.wikimedia.org/wiki/File:UMN_vs_LMN.png
10.6 https://commons.wikimedia.org/wiki/File:Sensory_and_motor_homunculi.jpg https://commons.wikimedia.org/wiki/File:Figure_35_03_04.jpg
10.7 https://commons.wikimedia.org/wiki/File:Intracranial_electrode_grid_for_electrocorticography.png
10.8 https://commons.wikimedia.org/wiki/File:Posterior_lobe_of_cerebellum_–_04.png
10.9 https://commons.wikimedia.org/wiki/File:CajalCerebellum.jpg
10.10 ttps://commons.wikimedia.org/wiki/File:Gray705.png
10.11 https://commons.wikimedia.org/wiki/File:CerebellumDiv.png modified by Austin Lim https://commons.wikimedia.org/ wiki/File:1613_Major_Regions_of_the_Cerebellum-02.jpg modified by Austin Lim
10.12 ttps://commons.wikimedia.org/wiki/File:CerebellumDiv.png modified by Austin Lim
10.13 https://commons.wikimedia.org/wiki/File:Brain_chrischan.jpg https://commons.wikimedia.org/wiki/File:Ataxia.jpg
10.14 Consalez GG, Goldowitz D, Casoni F and Hawkes R (2021) Origins, Development, and Compartmentation of the Granule Cells of the Cerebellum. Front. Neural Circuits 14:611841. doi: 10.3389/fncir.2020.611841
10.15 https://commons.wikimedia.org/wiki/File:Basal-ganglia-2.jpg modified by Austin Lim
10.16 https://commons.wikimedia.org/wiki/File:Confocal_image_of_spiny_neuron_-_1.jpg
10.17 https://commons.wikimedia.org/wiki/File:Skinner%27s_box.jpg
10.18 https://commons.wikimedia.org/wiki/File:Huntington%27s_disease_(5880985560).jpg modified by Austin Lim
10.19 https://commons.wikimedia.org/wiki/File:Typical_deep_brain_stimulation_setup.jpg modified by Austin Lim https://commons.wikimedia.org/wiki/File:Deep_brain_stimulation_electrode_placement_reconstruction.png
10.20 https://en.wikipedia.org/wiki/Pathophysiology_of_Parkinson%27s_disease#/media/File:Blausen_0704_ ParkinsonsDisease.png Blausen.com staff (2014). “Medical gallery of Blausen Medical 2014”. WikiJournal of Medicine 1 (2). DOI:10.15347/wjm/2014.010. ISSN 2002-4436. – Own work https://commons.wikimedia.org/wiki/File:Lewy_body_in_the_substantia_nigra_from_a_person_with_Parkinson%27s_ disease.jpg
10.21 https://commons.wikimedia.org/wiki/File:Pyramid_(medulla_oblongata)_-_lateral_view.png https://commons. wikimedia.org/wiki/File:Gray679.png
10.22 https://commons.wikimedia.org/wiki/File:Motor_Facial_Route.png Patrick J. Lynch; illustrator; C. Carl Jaffe; MD; cardiologist Yale University Center for Advanced Instructional Media Medical Illustrations by Patrick Lynch, generated for multimedia teaching projects by the Yale University School of Medicine, Center for Advanced Instructional Media, 1987- 2000. Patrick J. Lynch, http://patricklynch.net. Modified by Austin Lim
10.23 https://commons.wikimedia.org/wiki/File:1426_Corticospinal_Pathway.jpg modified by Austin Lim
10.24 ttps://commons.wikimedia.org/wiki/File:Cpg_output.jpg
10.25 Espinal A, Rostro-Gonzalez H, Carpio M, Guerra-Hernandez EI, Ornelas-Rodriguez M and Sotelo-Figueroa M (2016) Design of Spiking Central Pattern Generators for Multiple Locomotion Gaits in Hexapod Robots by Christiansen Grammar Evolution. Front. Neurorobot. 10:6. doi: 10.3389/fnbot.2016.00006
10.26 https://commons.wikimedia.org/wiki/File:Electron_micrograph_of_neuromuscular_junction_(cross-section).jpg https://commons.wikimedia.org/wiki/File:The_Muscle_Contraction_Process.png Modified by Austin Lim
10. 27 https://commons.wikimedia.org/wiki/File:Myasthenia_Gravis.jpg
10.28 https://commons.wikimedia.org/wiki/File:Sarcomere_relaxed_contracted.PNG
10.29 https://commons.wikimedia.org/wiki/File:Skeletal_muscle_400x.jpghttps://commons.wikimedia.org/wiki/File:Cardiac_ muscle_histology_400x.jpg
https://commons.wikimedia.org/wiki/File:Smooth_muscle_400x.jpg
10.30 https://commons.wikimedia.org/wiki/File:Animation_triceps_biceps.gif Modified by Austin Lim
